BACTERIAS
BACTERIAS
Tipos de bacterias
Las bacterias son microorganismos unicelulares procariontes, pertenecientes al mundo de las móneras y son los causantes de enfermedades, putrefacciones y fermentaciones en seres vivos y materias orgánicas.
Existen distintos criterios para clasificar a las bacterias, algunos de ellos son:
1) Según la respiración:
Bacterias aerobias:
Para respirar, este tipo de bacterias se valen del oxígeno.
Bacterias anaerobias:
estas, en cambio, no utilizan oxígeno, sino que deben sustituirlo por moléculas inorgánicas como las del sulfato o carbonato.
2) Según sus necesidades de crecimiento:
Bacterias autótrofas:
Estas bacterias tienen la capacidad de sintetizar las sustancias que necesitan para su metabolismo de sustancias inorgánicas. Dentro de este tipo se encuentran las fotosintetizantes que, gracias a los pigmentos que las componen, se valen de la energía de las radiaciones luminosas. También existen las quimiosintetizantes que utilizan la energía generada a partir de las reacciones químicas generadas por la oxidación.
Bacterias heterótrofas:
Este tipo de bacterias parasitan a los seres vivos y usan los compuestos orgánicos que estos elaboran. Dentro de este grupo existen las bacterias patógenas o parásitarias son las causantes de enfermedades en los seres vivos. También están aquellas bacterias de la putrefacción o saprófitas que descomponen las sustancias orgánicas en las que viven y se valen de su materia orgánica muerta para poder alimentarse. Otro tipo de bacterias son las simbióticas, que viven en cooperación con otros organismos. Por último están aquellas que realizan fermentaciones, de las que se vale el humano, como el ácido acético, fermentos lácticos, entre otros.
3) Según su forma:
A) Cocos:
Estas, también conocidas como bacterias redondeadas, pueden hallarse de forma aislada, como los micrococos,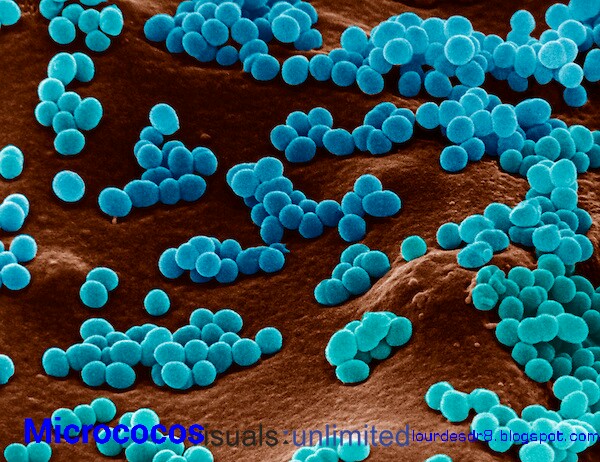 en pares, como los diplococos,
en pares, como los diplococos,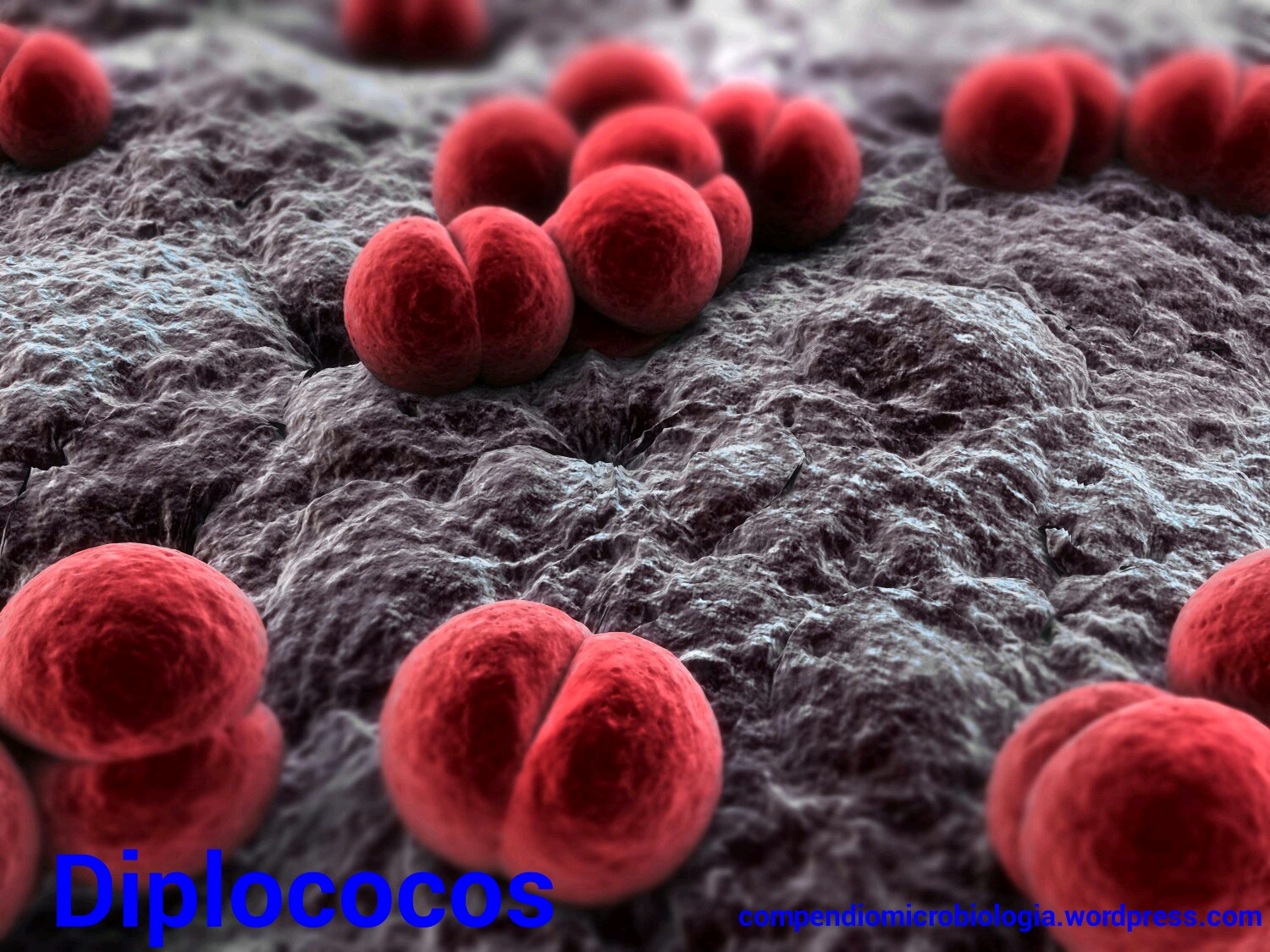 en cadena arracimada, como es el caso de los estafilcocos
en cadena arracimada, como es el caso de los estafilcocos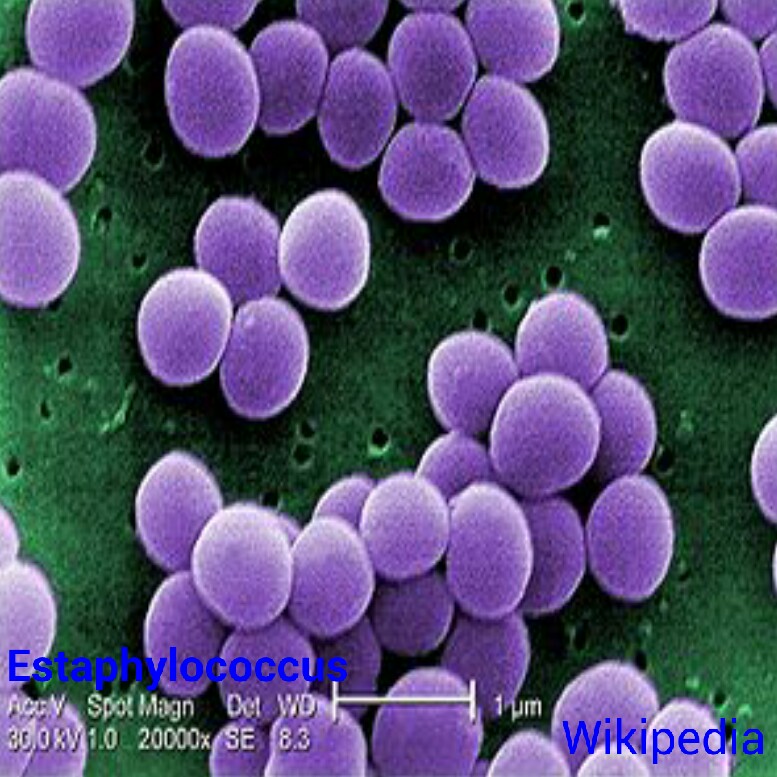 o, en cadenas arrosariadas como los estreptococos.
o, en cadenas arrosariadas como los estreptococos.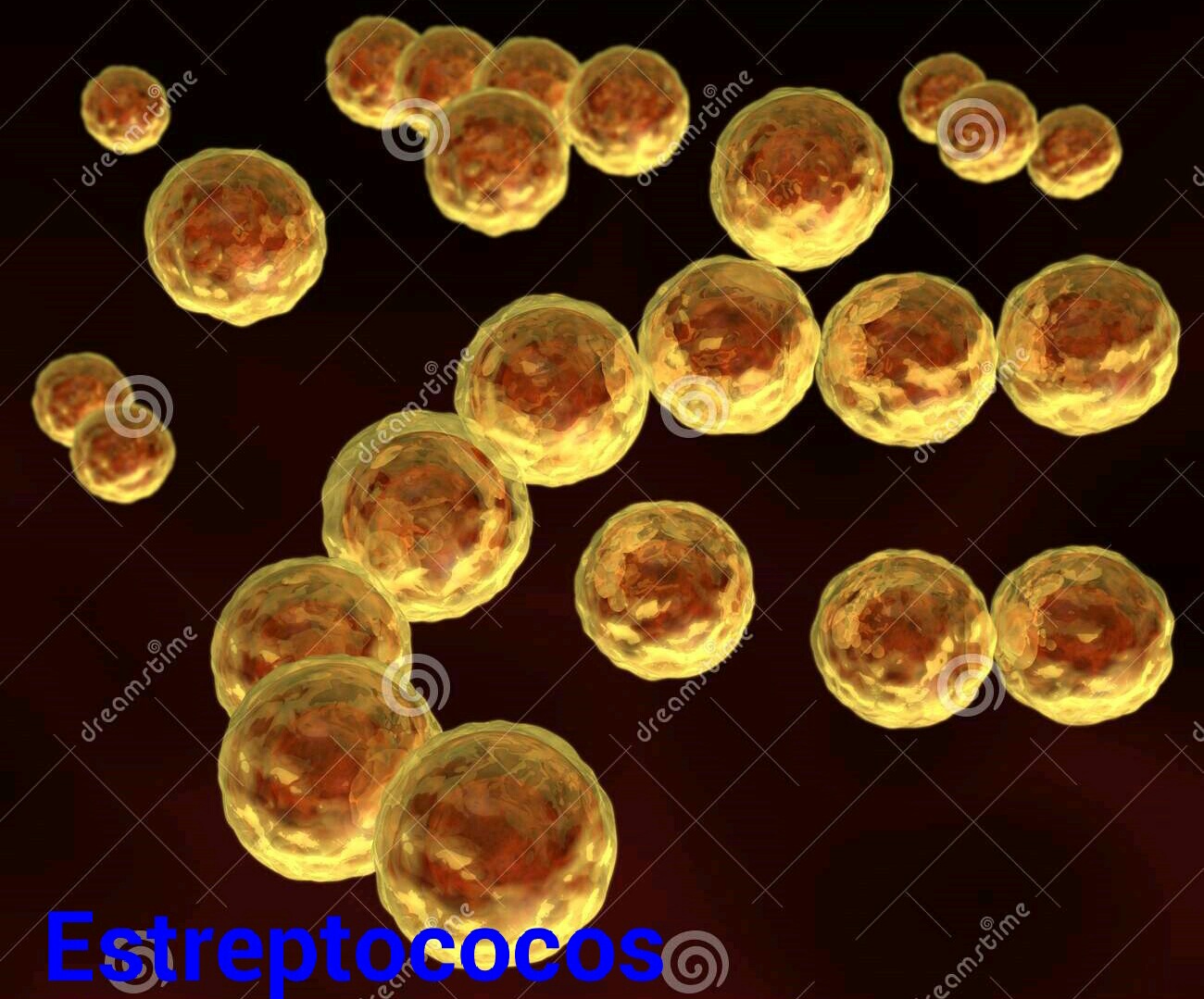
B) Leptothrix:
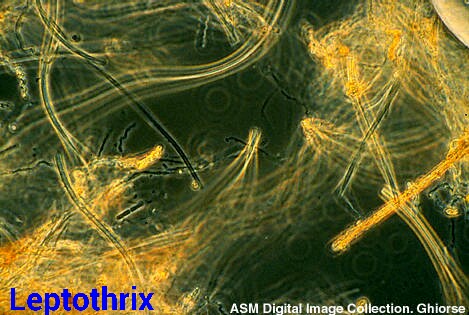
Son bacterias que las conforman filamentos tabicados y su tamaño es grande.
C) Bacilos: 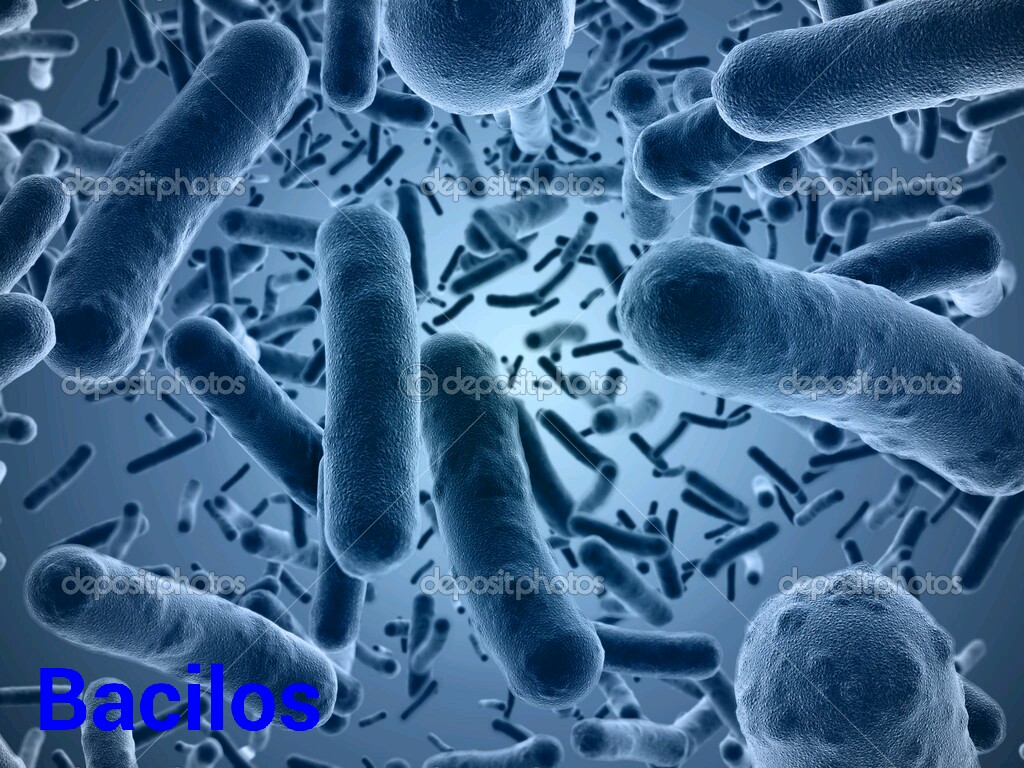
Son alargadas, curvas o rectas y pueden o no poseer flagelos.
D) Espirilos:
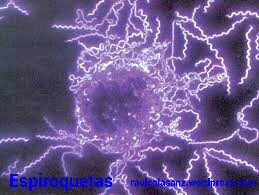
Estas bacterias son curvas helicoidalmente y pueden poseer un arrollamiento completo, como es el caso de las espiroquetas, o bien, un arrollamiento incompleto, como el de los vibriones.
Fuente: tiposde.org
Acá más información de Wikipedia :
Bacteria

Las bacterias son microorganismos procariotas que presentan un tamaño de unos pocos micrómetros (por lo general entre 0,5 y 5 μm de longitud) y diversas formas incluyendo filamentos, esferas (cocos), barras (bacilos), sacacorchos (vibrios) y hélices (espirilos). Las bacterias son células procariotas, por lo que a diferencia de las células eucariotas (de animales,plantas, hongos, etc.), no tienen el núcleo definido ni presentan, en general, orgánulos membranososinternos. Generalmente poseen una pared celular y ésta se compone de peptidoglicano. Muchas bacterias disponen de flagelos o de otros sistemas de desplazamiento y son móviles. Del estudio de las bacterias se encarga la bacteriología, una rama de lamicrobiología. La presencia frecuente de pared de peptidoglicano junto con su composición en lípidos de membrana son la principal diferencia que presentan frente a las arqueas, el otro importante grupo de microorganismos procariotas.
Las bacterias son los organismos más abundantes del planeta. Son ubicuas, se encuentran en todos loshábitats terrestres y acuáticos; crecen hasta en los más extremos como en los manantiales de aguas calientes y ácidas, en desechos radioactivos,[3] en las profundidades tanto del mar como de la corteza terrestre. Algunas bacterias pueden incluso sobrevivir en las condiciones extremas del espacio exterior. Se estima que se pueden encontrar en torno a 40 millones de células bacterianas en un gramo de tierra y un millón de células bacterianas en un mililitro de agua dulce. En total, se calcula que hay aproximadamente 5×1030bacterias en el mundo.[4]
Las bacterias son imprescindibles para el reciclaje de los elementos, pues muchos pasos importantes de losciclos biogeoquímicos dependen de éstas. Como ejemplo cabe citar la fijación del nitrógeno atmosférico. Sin embargo, solamente la mitad de los filos conocidos de bacterias tienen especies que se pueden cultivar en el laboratorio,[5] por lo que una gran parte (se supone que cerca del 90 %) de las especies de bacterias existentes todavía no ha sido descrita.
En el cuerpo humano hay aproximadamente diez veces tantas células bacterianas como células humanas, con una gran cantidad de bacterias en la piel y en el tracto digestivo.[6] Aunque el efecto protector del sistema inmunológico hace que la gran mayoría de estas bacterias sea inofensiva o beneficiosa, algunas bacterias patógenas pueden causar enfermedades infecciosas, incluyendo cólera, difteria, escarlatina,lepra, sífilis, tifus, etc. Las enfermedades bacterianas mortales más comunes son las infecciones respiratorias, con una mortalidad solo para latuberculosis de cerca de dos millones de personas al año.[7]
En todo el mundo se utilizan antibióticos para tratar las infecciones bacterianas. Los antibióticos son efectivos contra las bacterias ya que inhiben la formación de la pared celular o detienen otros procesos de su ciclo de vida. También se usan extensamente en la agricultura y la ganadería en ausencia de enfermedad, lo que ocasiona que se esté generalizando la resistencia de las bacterias a los antibióticos. En la industria, las bacterias son importantes en procesos tales como el tratamiento de aguas residuales, en la producción demantequilla, queso, vinagre, yogur, etc., y en la fabricación de medicamentos y de otros productos químicos.[8]
Aunque el término bacteria incluía tradicionalmente a todos los procariotas, actualmente la taxonomía y la nomenclatura científica los divide en dos grupos. Estos dominios evolutivos se denominan Bacteria y Archaea (arqueas).[9] La división se justifica en las grandes diferencias que presentan ambos grupos a nivel bioquímico y genético.
Origen y evolución de las bacterias Editar
Árbol filogenético mostrando la divergencia de las especies modernas de su ancestro común en el centro.[27] Los tres dominios están coloreados de la siguiente forma; las bacterias en azul, las archaeas en verde, y las eucariotas de color rojo.
Los seres vivos se dividen actualmente en tres dominios: bacterias (Bacteria), arqueas (Archaea) y eucariontes (Eukarya). En los dominios Archaea y Bacteria se incluyen los organismos procariotas, esto es, aquellos cuyas células no tienen un núcleo celular diferenciado, mientras que en el dominio Eukarya se incluyen las formas de vida más conocidas y complejas (protistas, animales, hongos y plantas).
El término "bacteria" se aplicó tradicionalmente a todos los microorganismos procariotas. Sin embargo, la filogenia molecular ha podido demostrar que los microorganismos procariotas se dividen en dos dominios, originalmente denominados Eubacteria y Archaebacteria, y ahora renombrados como Bacteria y Archaea,[28] que evolucionaron independientemente desde un ancestro común. Estos dos dominios, junto con el dominio Eukarya, constituyen la base del sistema de tres dominios, que actualmente es el sistema de clasificación más ampliamente utilizado en bacteriología.[29]
El término Mónera, actualmente en desuso, en la antigua clasificación de los cinco reinos significaba lo mismo que procariota, y así sigue siendo usado en muchos manuales y libros de texto.
Los antepasados de los procariotas modernos fueron los primeros organismos (las primeras células) que se desarrollaron sobre la tierra, hace unos 3.800-4.000 millones años. Durante cerca de 3.000 millones de años más, todos los organismos siguieron siendo microscópicos, siendo probablemente bacterias y arqueas las formas de vida dominantes.[30] [31] Aunque existen fósiles bacterianos, por ejemplo los estromatolitos, al no conservar su morfología distintiva no se pueden emplear para estudiar la historia de la evolución bacteriana, o el origen de una especie bacteriana en particular. Sin embargo, las secuencias genéticas sí se pueden utilizar para reconstruir la filogenia de los seres vivos, y estos estudios sugieren que arqueas y eucariontes están más relacionados entre sí que con las bacterias.[32]
En la actualidad se discute si los primeros procariotas fueron bacterias o arqueas. Algunos investigadores piensan que Bacteria es el dominio más antiguo con Archaea y Eukarya derivando a partir de él,[29] mientras que otros consideran que el dominio más antiguo es Archaea.[33] Se ha propuesto que el ancestro común más reciente de bacterias y arqueas podría ser un hipertermófilo que vivió entre 2.500 y 3.200 millones de años atrás.[34] [35] En cambio, otros científicos sostienen que tanto Archaea como Eukarya son relativamente recientes (de hace unos 900 millones de años)[36] [37] y que evolucionaron a partir de una bacteria Gram-positiva (probablemente una Actinobacteria), que mediante la sustitución de la pared bacteriana de peptidoglicano por otra de glicoproteína daría lugar a un organismo Neomura.[38] [39]
Las bacterias también han estado implicadas en la segunda gran divergencia evolutiva, la que separó Archaea de Eukarya. Se considera que las mitocondrias de los eucariontes proceden de la endosimbiosis de una proteobacteria alfa.[40] [41] En este caso, el antepasado de los eucariontes, que posiblemente estaba relacionado con las arqueas (el organismo Neomura), ingirió una proteobacteria que, al escapar a la digestión, se desarrolló en el citoplasma y dio lugar a las mitocondrias. Éstas se pueden encontrar en todos los eucariontes, aunque a veces en formas muy reducidas, como en los protistas amitocondriales. Después, e independientemente, una segunda endosimbiosis por parte de algún eucarionte mitocondrial con una cianobacteria condujo a la formación de los cloroplastos de algas y plantas. Se conocen incluso algunos grupos de algas que se han originado claramente de acontecimientos posteriores de endosimbiosis por parte de eucariotas heterótrofos que, tras ingerir algas eucariotas, se convirtieron en plastos de segunda generación.[42] [43]
Morfología bacteriana Editar
Existen bacterias con múltiples morfologías.
Las bacterias presentan una amplia variedad de tamaños y formas. La mayoría presentan un tamaño diez veces menor que el de las células eucariotas, es decir, entre 0,5 y 5 μm. Sin embargo, algunas especies como Thiomargarita namibiensis y Epulopiscium fishelsoni llegan a alcanzar los 0,5 mm, lo cual las hace visibles al ojo desnudo.[44] En el otro extremo se encuentran bacterias más pequeñas conocidas, entre las que cabe destacar las pertenecientes al género Mycoplasma, las cuales llegan a medir solo 0,3 μm, es decir, tan pequeñas como los virus más grandes.[45]
La forma de las bacterias es muy variada y, a menudo, una misma especie adopta distintos tipos morfológicos, lo que se conoce como pleomorfismo. De todas formas, podemos distinguir tres tipos fundamentales de bacterias:
Coco (del griego kókkos, grano): de forma esférica.
Diplococo: cocos en grupos de dos.
Tetracoco: cocos en grupos de cuatro.
Estreptococo: cocos en cadenas.
Estafilococo: cocos en agrupaciones irregulares o en racimo.
Bacilo (del latín baculus, varilla): en forma de bastoncillo.
Formas helicoidales:
Vibrio: ligeramente curvados y en forma de coma, judía o cacahuete.
Espirilo: en forma helicoidal rígida o en forma de tirabuzón.
Espiroqueta: en forma de tirabuzón (helicoidal flexible).
Algunas especies presentan incluso formas tetraédricas o cúbicas.[46] Esta amplia variedad de formas es determinada en última instancia por la composición de la pared celular y el citoesqueleto, siendo de vital importancia, ya que puede influir en la capacidad de la bacteria para adquirir nutrientes, unirse a superficies o moverse en presencia de estímulos.[47] [48]
A continuación se citan diferentes especies con diversos patrones de asociación:
Neisseria gonorrhoeae en forma diploide (por pares).
Streptococcus en forma de cadenas.
Staphylococcus en forma de racimos.
Actinobacteria en forma de filamentos. Dichos filamentos suelen rodearse de una vaina que contiene multitud de células individuales, pudiendo llegar a ramificarse, como el género Nocardia, adquiriendo así el aspecto del micelio de un hongo.[49]
Rango de tamaños que presentan las células procariotas en relación a otros organismos y biomoléculas.
Las bacterias presentan la capacidad de anclarse a determinadas superficies y formar un agregado celular en forma de capa denominado biopelícula o biofilme, los cuales pueden tener un grosor que va desde unos pocos micrómetros hasta medio metro. Estas biopelículas pueden congregar diversas especies bacterianas, además de protistas y arqueas, y se caracterizan por formar un conglomerado de células y componentes extracelulares, alcanzando así un nivel mayor de organización o estructura secundaria denominada microcolonia, a través de la cual existen multitud de canales que facilitan la difusión de nutrientes.[50] [51] En ambientes naturales tales como el suelo o la superficie de las plantas, la mayor parte de las bacterias se encuentran ancladas a las superficies en forma de biopelículas.[52] Dichas biopelículas deben ser tenidas en cuenta en las infecciones bacterianas crónicas y en los implantes médicos, ya que las bacterias que forman estas estructuras son mucho más difíciles de erradicar que las bacterias individuales.[53]
Por último, cabe destacar un tipo de morfología más compleja aún, observable en algunos microorganismos del grupo de las mixobacterias. Cuando estas bacterias se encuentran en un medio escaso en aminoácidos son capaces de detectar a las células de alrededor, en un proceso conocido como percepción de quórum, en el cual todas las células migran hacia las demás y se agregan, dando lugar a cuerpos fructíferos que pueden alcanzar los 0,5 mm de longitud y contener unas 100.000 células.[54] Una vez formada dicha estructura las bacterias son capaces de llevar a cabo diferentes funciones, es decir, se diferencian, alcanzando así un cierto nivel de organización pluricelular. Por ejemplo, entre una y diez células migran a la parte superior del cuerpo fructífero y, una vez allí, se diferencian para dar lugar a un tipo de células latentes denominadas mixosporas, las cuales son más resistentes a la desecación y, en general, a condiciones ambientales adversas.[55]
Estructura de la célula bacteriana Editar
Estructura de la célula bacteriana. A-Pili; B-Ribosomas; C-Cápsula; D-Pared celular; E-Flagelo; F-Citoplasma; G-Vacuola; H-Plásmido; I-Nucleoide; J-Membrana citoplasmática.
Las bacterias son organismos relativamente sencillos. Sus dimensiones son muy reducidas, unos 2 μm de ancho por 7-8 μm de longitud en la forma cilíndrica (bacilo) de tamaño medio; aunque son muy frecuentes las especies de 0,5-1,5 μm.
Al tratarse de organismos procariotas, tienen las características básicas correspondientes como la carencia de un núcleo delimitado por una membrana aunque presentan un nucleoide, una estructura elemental que contiene una gran molécula circular de ADN. El citoplasma carece de orgánulos delimitados por membranas y de las formaciones protoplasmáticas propias de las células eucariotas. En el citoplasma se pueden apreciar plásmidos, pequeñas moléculas circulares de ADN que coexisten con el nucleoide, contienen genes y son comúnmente usados por los procariontes en la conjugación. El citoplasma también contiene vacuolas (gránulos que contienen sustancias de reserva) y ribosomas (utilizados en la síntesis de proteínas).
Una membrana citoplasmática compuesta de lípidos rodea el citoplasma y, al igual que las células de las plantas, la mayoría posee una pared celular, que en este caso está compuesta por peptidoglicano (mureína). La mayoría de bacterias, presentan además una segunda membrana lipídica (membrana externa) rodeando a la pared celular. El espacio comprendido entre la membrana citoplasmática y la pared celular (o la membrana externa si esta existe) se denomina espacio periplásmico. Algunas bacterias presentan una cápsula y otras son capaces de desarrollarse como endosporas, estados latentes capaces de resistir condiciones extremas. Entre las formaciones exteriores propias de la célula bacteriana destacan los flagelos y los pili.
Estructuras intracelulares Editar
La membrana citoplasmática de las bacterias es similar a la de plantas y animales, si bien generalmente no presenta colesterol.
La membrana citoplasmática bacteriana tiene una estructura similar a la de plantas y animales. Es una bicapa lipídica compuesta fundamentalmente de fosfolípidos en la que se insertan moléculas de proteínas. En las bacterias realiza numerosas funciones entre las que se incluyen las de barrera osmótica, transporte, biosíntesis, transducción de energía, centro de replicación de ADN y punto de anclaje para los flagelos. A diferencia de las membranas eucarióticas, generalmente no contiene esteroles (son excepciones micoplasmas y algunas proteobacterias), aunque puede contener componentes similares denominados hopanoides.
Muchas importantes reacciones bioquímicas que tienen lugar en las células se producen por la existencia de gradientes de concentración a ambos lados de una membrana. Este gradiente crea una diferencia de potencial análoga a la de una batería eléctrica y permite a la célula, por ejemplo, el transporte de electrones y la obtención de energía. La ausencia de membranas internas en las bacterias significa que estas reacciones tienen que producirse a través de la propia membrana citoplasmática, entre el citoplasma y el espacio periplásmico.[56]
Puesto que las bacterias son procariotas no tienen orgánulos citoplasmáticos delimitados por membranas y por ello presentan pocas estructuras intracelulares. Carecen de núcleo celular, mitocondrias, cloroplastos y de los otros orgánulos presentes en las células eucariotas, tales como el aparato de Golgi y el retículo endoplasmático.[57] Como excepción, algunas bacterias contienen estructuras intracelulares rodeadas por membranas que pueden considerarse primitivos orgánulos. Ejemplos son los tilacoides de las cianobacterias, los compartimentos que contienen amonio monooxigenasa en Nitrosomonadaceae y diversas estructuras en Planctomycetes.[58]
Como todos los organismos vivos, las bacterias contienen ribosomas para la síntesis de proteínas, pero éstos son diferentes a los de eucariotas.[59] La estructura de los ribosomas y el ARN ribosomal de arqueas y bacterias son similares, ambos ribosomas son de tipo 70S mientras que los ribosomas eucariotas son de tipo 80S. Sin embargo, la mayoría de las proteínas ribosomiales, factores de traducción y ARNt arqueanos son más parecidos a los eucarióticos que a los bacterianos.
Muchas bacterias presentan vacuolas, gránulos intracelulares para el almacenaje de sustancias, como por ejemplo glucógeno,[60] polifosfatos,[61] azufre[62] o polihidroxialcanoatos.[63] Ciertas especies bacterianas fotosintéticas, tales como las cianobacterias, producen vesículas internas de gas que utilizan para regular su flotabilidad y así alcanzar la profundidad con intensidad de luz óptima o unos niveles de nutrientes óptimos.[64] Otras estructuras presentes en ciertas especies son los carboxisomas (que contienen enzimas para la fijación de carbono) y los magnetosomas (para la orientación magnética).
Elementos del citoesqueleto de Caulobacter crescentus. En la figura, estos elementos procarióticos se relacionan con sus homólogos eucariotas y se hipotetiza su función celular.[65] Debe tenerse en cuenta que las funciones en la pareja FtsZ-MreB se invirtieron durante la evolución al convertirse en tubulina-actina.
Las bacterias no tienen un núcleo delimitado por membranas. El material genético está organizado en un único cromosoma situado en el citoplasma, dentro de un cuerpo irregular denominado nucleoide.[66] La mayoría de los cromosomas bacterianos son circulares, si bien existen algunos ejemplos de cromosomas lineales, por ejemplo, Borrelia burgdorferi. El nucleoide contiene el cromosoma junto con las proteínas asociadas y ARN. El orden Planctomycetes es una excepción, pues una membrana rodea su nucleoide y tiene varias estructuras celulares delimitadas por membranas.[58]
Anteriormente se pensaba que las células procariotas no poseían citoesqueleto, pero desde entonces se han encontrado homólogos bacterianos de las principales proteínas del citoesqueleto de los eucariontes.[67] Estos incluyen las proteínas estructurales FtsZ (que se ensambla en un anillo para mediar durante la división celular bacteriana) y MreB (que determina la anchura de la célula). El citoesqueleto bacteriano desempeña funciones esenciales en la protección, determinación de la forma de la célula bacteriana y en la división celular.[68]
Estructuras extracelulares Editar
Las bacterias disponen de una pared celular que rodea a su membrana citoplasmática. Las paredes celulares bacterianas están hechas de peptidoglicano (llamado antiguamente mureína). Esta sustancia está compuesta por cadenas de polisacárido enlazadas por péptidos inusuales que contienen aminoácidos D.[69] Estos aminoácidos no se encuentran en las proteínas, por lo que protegen a la pared de la mayoría de las peptidasas. Las paredes celulares bacterianas son distintas de las que tienen plantas y hongos, compuestas de celulosa y quitina, respectivamente.[70] Son también distintas a las paredes celulares de Archaea, que no contienen peptidoglicano. El antibiótico penicilina puede matar a muchas bacterias inhibiendo un paso de la síntesis del peptidoglicano.[70]
Paredes celulares bacterianas. Arriba: Bacteria Gram positiva. 1-membrana citoplasmática, 2-pared celular, 3-espacio periplásmico. Abajo: Bacteria Gram negativa. 4-membrana citoplasmática, 5-pared celular, 6-membrana externa, 7-espacio periplásmico.
Existen dos diferentes tipos de pared celular bacteriana denominadas Gram-positiva y Gram-negativa, respectivamente. Estos nombres provienen de la reacción de la pared celular a la tinción de Gram, un método tradicionalmente empleado para la clasificación de las especies bacterianas.[71] Las bacterias Gram-positivas tienen una pared celular gruesa que contiene numerosas capas de peptidoglicano en las que se inserta ácido teicoico. En cambio, las bacterias Gram-negativas tienen una pared relativamente fina, consistente en unas pocas capas de peptidoglicano, rodeada por una segunda membrana lipídica (la membrana externa) que contiene lipopolisacáridos y lipoproteínas.
Las micoplasmas son una excepción, pues carecen de pared celular. La mayoría de las bacterias tienen paredes celulares Gram-negativas; solamente son Gram-positivas Firmicutes y Actinobacteria. Estos dos grupos eran antiguamente conocidos como bacterias Gram-positivas de contenido GC bajo y bacterias Gram-positivas de contenido GC alto, respectivamente.[72] Estas diferencias en la estructura de la pared celular dan lugar a diferencias en la susceptibilidad antibiótica. Por ejemplo, la vancomicina puede matar solamente a bacterias Gram-positivas y es ineficaz contra patógenos Gram-negativos, tales como Haemophilus influenzae o Pseudomonas aeruginosa.[73] Dentro del filo Actinobacteria cabe hacer una mención especial al género Mycobacterium, el cual, si bien se encuadra dentro de las Gram positivas, no parece serlo desde el punto de vista empírico, ya que su pared no retiene el tinte. Esto se debe a que presentan una pared celular poco común, rica en ácidos micólicos, de carácter hidrófobo y ceroso y bastante gruesa, lo que les confiere una gran resistencia.
Helicobacter pylori visto al microscopio electrónico, mostrando numerosos flagelos sobre la superficie celular.
Muchas bacterias tienen una capa S de moléculas de proteína de estructura rígida que cubre la pared celular.[74] Esta capa proporciona protección química y física para la superficie celular y puede actuar como una barrera de difusión macromolecular. Las capas S tienen diversas (aunque todavía no bien comprendidas) funciones. Por ejemplo, en el género Campylobacter actúan como factores de virulencia y en la especie Bacillus stearothermophilus contienen enzimas superficiales.[75]
Los flagelos son largos apéndices filamentosos compuestos de proteínas y utilizados para el movimiento. Tienen un diámetro aproximado de 20 nm y una longitud de hasta 20 μm. Los flagelos son impulsados por la energía obtenida de la transferencia de iones. Esta transferencia es impulsada por el gradiente electroquímico que existe entre ambos lados de la membrana citoplasmática.[76]
Escherichia coli presenta unas 100-200 fimbrias que utiliza para adherirse a las células epiteliales o al tracto urogenital.
Las fimbrias son filamentos finos de proteínas que se distribuyen sobre la superficie de la célula. Tienen un diámetro aproximado de 2-10 nm y una longitud de hasta varios μm. Cuando se observan a través del microscopio electrónico se asemejan a pelos finos. Las fimbrias ayudan a la adherencia de las bacterias a las superficies sólidas o a otras células y son esenciales en la virulencia de algunos patógenos.[77] Los pili son apéndices celulares ligeramente mayores que las fimbrias y se utilizan para la transferencia de material genético entre bacterias en un proceso denominado conjugación bacteriana.[78]
Estructuras extracelulares bacterianas: 1-cápsula, 2-glicocalix (capa mucosa), 3-biopelícula.
Muchas bacterias son capaces de acumular material en el exterior para recubrir su superficie. Dependiendo de la rigidez y su relación con la célula se clasifican en cápsulas y glicocalix. La cápsula es una estructura rígida que se une firmemente a la superficie bacteriana, en tanto que el glicocalix es flexible y se une de forma laxa. Estas estructuras protegen a las bacterias pues dificultan que sean fagocitadas por células eucariotas tales como los macrófagos.[79] También pueden actuar como antígenos y estar implicadas en el reconocimiento bacteriano, así como ayudar a la adherencia superficial y a la formación de biopelículas.[80]
La formación de estas estructuras extracelulares depende del sistema de secreción bacteriano. Este sistema transfiere proteínas desde el citoplasma al periplasma o al espacio que rodea a la célula. Se conocen muchos tipos de sistemas de secreción, que son a menudo esenciales para la virulencia de los patógenos, por lo que son extensamente estudiados.[81]
Endosporas Editar
Véase también: Esporas bacterianas
Bacillus anthracis (teñido púrpura) desarrollándose en el líquido cefalorraquídeo. Cada pequeño segmento es una bacteria.
Ciertos géneros de bacterias Gram-positivas, tales como Bacillus, Clostridium, Sporohalobacter, Anaerobacter y Heliobacterium, pueden formar endosporas.[82] Las endosporas son estructuras durmientes altamente resistentes cuya función primaria es sobrevivir cuando las condiciones ambientales son adversas. En casi todos los casos, las endosporas no forman parte de un proceso reproductivo, aunque Anaerobacter puede formar hasta siete endosporas a partir de una célula.[83] Las endosporas tienen una base central de citoplasma que contiene ADN y ribosomas, rodeada por una corteza y protegida por una cubierta impermeable y rígida.
Las endosporas no presentan un metabolismo detectable y pueden sobrevivir a condiciones físicas y químicas extremas, tales como altos niveles de luz ultravioleta, rayos gamma, detergentes, desinfectantes, calor, presión y desecación.[84] En este estado durmiente, las bacterias pueden seguir viviendo durante millones de años,[85] [86] e incluso pueden sobrevivir en la radiación y vacío del espacio exterior.[87] Las endosporas pueden también causar enfermedades. Por ejemplo, puede contraerse carbunco por la inhalación de endosporas de Bacillus anthracis y tétanos por la contaminación de las heridas con endosporas de Clostridium tetani.[88]
Metabolismo Editar
Artículo principal: Metabolismo microbiano
Filamento (una colonia) de cianobacteria fotosintética.
En contraste con los organismos superiores, las bacterias exhiben una gran variedad de tipos metabólicos.[89] La distribución de estos tipos metabólicos dentro de un grupo de bacterias se ha utilizado tradicionalmente para definir su taxonomía, pero estos rasgos no corresponden a menudo con las clasificaciones genéticas modernas.[90] El metabolismo bacteriano se clasifica con base en tres criterios importantes: el origen del carbono, la fuente de energía y los donadores de electrones. Un criterio adicional para clasificar a los microorganismos que respiran es el receptor de electrones usado en la respiración.[91]
Según la fuente de carbono, las bacterias se pueden clasificar como:
Heterótrofas, cuando usan compuestos orgánicos.
Autótrofas, cuando el carbono celular se obtiene mediante la fijación del dióxido de carbono.
Las bacterias autótrofas típicas son las cianobacterias fotosintéticas, las bacterias verdes del azufre y algunas bacterias púrpura. Pero hay también muchas otras especies quimiolitotrofas, por ejemplo, las bacterias nitrificantes y oxidantes del azufre.[92]
Según la fuente de energía, las bacterias pueden ser:
Fototrofas, cuando emplean la luz a través de la fotosíntesis.
Quimiotrofas, cuando obtienen energía a partir de sustancias químicas que son oxidadas principalmente a expensas del oxígeno (respiración aerobia) o de otros receptores de electrones alternativos (respiración anaerobia).
Según los donadores de electrones, las bacterias también se pueden clasificar como:
Litotrofas, si utilizan como donadores de electrones compuestos inorgánicos.
Organotrofas, si utilizan como donadores de electrones compuestos orgánicos.
Los organismos quimiotrofos usan donadores de electrones para la conservación de energía (durante la respiración aerobia, anaerobia y la fermentación) y para las reacciones biosintéticas (por ejemplo, para la fijación del dióxido de carbono), mientras que los organismos fototrofos los utilizan únicamente con propósitos biosintéticos.
Bacterias del hierro en un regato. Estos microorganismos quimiolitotrofos obtienen la energía que necesitan por oxidación del óxido ferroso a óxido férrico.
Los organismos que respiran usan compuestos químicos como fuente de energía, tomando electrones del sustrato reducido y transfiriéndolos a un receptor terminal de electrones en una reacción redox. Esta reacción desprende energía que se puede utilizar para sintetizar ATP y así mantener activo el metabolismo. En los organismos aerobios, el oxígeno se utiliza como receptor de electrones. En los organismos anaerobios se utilizan como receptores de electrones otros compuestos inorgánicos tales como nitratos, sulfatos o dióxido de carbono. Esto conduce a que se lleven a cabo los importantes procesos biogeoquímicos de la desnitrificación, la reducción del sulfato y la acetogénesis, respectivamente. Otra posibilidad es la fermentación, un proceso de oxidación incompleta, totalmente anaeróbico, siendo el producto final un compuesto orgánico, que al reducirse será el receptor final de los electrones. Ejemplos de productos de fermentación reducidos son el lactato (en la fermentación láctica), etanol (en la fermentación alcohólica), hidrógeno, butirato, etc. La fermentación es posible porque el contenido de energía de los sustratos es mayor que el de los productos, lo que permite que los organismos sinteticen ATP y mantengan activo su metabolismo.[93] [94] Los organismos anaerobios facultativos pueden elegir entre la fermentación y diversos receptores terminales de electrones dependiendo de las condiciones ambientales en las cuales se encuentren.
Las bacterias litotrofas pueden utilizar compuestos inorgánicos como fuente de energía. Los donadores de electrones inorgánicos más comunes son el hidrógeno, el monóxido de carbono, el amoníaco (que conduce a la nitrificación), el hierro ferroso y otros iones de metales reducidos, así como varios compuestos de azufre reducidos. En determinadas ocasiones, las bacterias metanotrofas pueden usar gas metano como fuente de electrones y como sustrato simultáneamente, para el anabolismo del carbono.[95] En la fototrofía y quimiolitotrofía aerobias, se utiliza el oxígeno como receptor terminal de electrones, mientras que bajo condiciones anaeróbicas se utilizan compuestos inorgánicos. La mayoría de los organismos litotrofos son autótrofos, mientras que los organismos organotrofos son heterótrofos.
Además de la fijación del dióxido de carbono mediante la fotosíntesis, algunas bacterias también fijan el gas nitrógeno usando la enzima nitrogenasa. Esta característica es muy importante a nivel ambiental y se puede encontrar en bacterias de casi todos los tipos metabólicos enumerados anteriormente, aunque no es universal.[96] El metabolismo microbiano puede jugar un papel importante en la biorremediación pues, por ejemplo, algunas especies pueden realizar el tratamiento de las aguas residuales y otras son capaces de degradar los hidrocarburos, sustancias tóxicas e incluso radiactivas. En cambio, las bacterias reductoras de sulfato son en gran parte responsables de la producción de formas altamente tóxicas de mercurio (metil- y dimetil-mercurio) en el ambiente.[97]
Movimiento Editar
Véase también: Flagelo bacteriano
Los diferentes tipos de disposición de los flagelos bacterianos: A-Monotrico; B-Lofotrico; C-Anfitrico; D-Peritrico.
Algunas bacterias son inmóviles y otras limitan su movimiento a cambios de profundidad. Por ejemplo, cianobacterias y bacterias verdes del azufre contienen vesículas de gas con las que pueden controlar su flotabilidad y así conseguir un óptimo de luz y alimento.[98] Las bacterias móviles pueden desplazarse por deslizamiento, mediante contracciones o más comúnmente usando flagelos. Algunas bacterias pueden deslizarse por superficies sólidas segregando una sustancia viscosa, pero el mecanismo que actúa como propulsor es todavía desconocido. En el movimiento mediante contracciones, la bacteria usa su pilus de tipo IV como gancho de ataque, primero lo extiende, anclándolo y después lo contrae con una fuerza notable (>80 pN).[99]
El flagelo bacteriano es un largo apéndice filamentoso helicoidal propulsado por un motor rotatorio (como una hélice) que puede girar en los dos sentidos. El motor utiliza como energía un gradiente electroquímico a través de la membrana. Los flagelos están compuestos por cerca de 20 proteínas, con aproximadamente otras 30 proteínas para su regulación y coordinación.[98] Hay que tener en cuenta que, dado el tamaño de la bacteria, el agua les resulta muy viscosa y el mecanismo de propulsión debe ser muy potente y eficiente. Los flagelos bacterianos se encuentran tanto en las bacterias Gram-positivas como Gram-negativas y son completamente diferentes de los eucarióticos y, aunque son superficialmente similares a los arqueanos, se consideran no homólogos.
El flagelo bacteriano es un apéndice movido por un motor rotatorio. El rotor puede girar a 6.000-17.000 rpm, pero el apéndice usualmente sólo alcanza 200-1000 rpm. 1-filamento, 2-espacio periplásmico, 3-codo, 4-juntura, 5-anillo L, 6-eje, 7-anillo P, 8-pared celular, 9-estátor, 10-anillo MS, 11-anillo C, 12-sistema de secreción de tipo III, 13-membrana externa, 14-membrana citoplasmática, 15-punta.
Según el número y disposición de los flagelos en la superficie de la bacteria se distinguen los siguientes tipos: un solo flagelo (monotrico), un flagelo en cada extremo (anfitrico), grupos de flagelos en uno o en los dos extremos (lofotrico) y flagelos distribuidos sobre toda la superficie de la célula (peritricos). En un grupo único de bacterias, las espiroquetas, se presentan unos flagelos especializados, denominados filamentos axiales, localizados intracelularmente en el espacio periplásmico, entre las dos membranas. Estos producen un movimiento rotatorio que hace que la bacteria gire como un sacacorchos desplazándose hacia delante.[98]
Muchas bacterias (tales como E. coli) tienen dos tipos de movimiento: en línea recta (carrera) y aleatorio. En este último, se realiza un movimiento tridimensional aleatorio al combinar la bacteria carreras cortas con virajes al azar.[100] Las bacterias móviles pueden presentar movimientos de atracción o repulsión determinados por diferentes estímulos. Estos comportamientos son denominados taxis, e incluyen diversos tipos como la quimiotaxis, la fototaxis o la magnetotaxis.[101] [102] En el peculiar grupo de las mixobacterias, las células individuales se mueven juntas formando ondas de células, que terminarán agregándose para formar los cuerpos fructíferos característicos de este género.[103] El movimiento de las mixobacterias se produce solamente sobre superficies sólidas, en contraste con E. coli, que es móvil tanto en medios líquidos como sólidos.
Varias especies de Listeria y Shigella se mueven dentro de las células huésped apropiándose de su citoesqueleto, que normalmente movería los orgánulos. La polimerización de actina crea un empuje en un extremo de la bacteria que la mueve a través del citoplasma de la célula huésped.[104]
Reproducción Editar
Modelo de divisiones binarias sucesivas en el microorganismo Escherichia coli.
En las bacterias, el aumento en el tamaño de las células (crecimiento) y la reproducción por división celular están íntimamente ligados, como en la mayor parte de los organismos unicelulares. Las bacterias crecen hasta un tamaño fijo y después se reproducen por fisión binaria, una forma de reproducción asexual.[105] En condiciones apropiadas, una bacteria Gram-positiva puede dividirse cada 20–30 minutos y una Gram-negativa cada 15–20 minutos, y en alrededor de 16 horas su número puede ascender a unos 5.000 millones (aproximadamente el número de personas que habitan la Tierra). Bajo condiciones óptimas, algunas bacterias pueden crecer y dividirse muy rápido, tanto como cada 9,8 minutos.[106] En la división celular se producen dos células hijas idénticas. Algunas bacterias, todavía reproduciéndose asexualmente, forman estructuras reproductivas más complejas que facilitan la dispersión de las células hijas recién formadas. Ejemplos incluyen la formación de cuerpos fructíferos (esporangios) en las mixobacterias, la formación de hifas en Streptomyces y la gemación. En la gemación una célula forma una protuberancia que a continuación se separa y produce una nueva célula hija.
Por otro lado, cabe destacar un tipo de reproducción sexual en bacterias, denominada parasexualidad bacteriana. En este caso, las bacterias son capaces de intercambiar material genético en un proceso conocido como conjugación bacteriana. Durante el proceso una bacteria donante y una bacteria receptora llevan a cabo un contacto mediante pelos sexuales huecos o pili, a través de los cuales se transfiere una pequeña cantidad de ADN independiente o plásmido conjugativo. El mejor conocido es el plásmido F de E. coli, que además puede integrarse en el cromosoma bacteriano. En este caso recibe el nombre de episoma, y en la transferencia arrastra parte del cromosoma bacteriano. Se requiere que exista síntesis de ADN para que se produzca la conjugación. La replicación se realiza al mismo tiempo que la transferencia.
Crecimiento Editar
Fases del crecimiento bacteriano.
El crecimiento bacteriano sigue tres fases. Cuando una población bacteriana se encuentra en un nuevo ambiente con elevada concentración de nutrientes que le permiten crecer necesita un período de adaptación a dicho ambiente. Esta primera fase se denomina fase de adaptación o fase lag y conlleva un lento crecimiento, donde las células se preparan para comenzar un rápido crecimiento, y una elevada tasa de biosíntesis de las proteínas necesarias para ello, como ribosomas, proteínas de membrana, etc.[107] La segunda fase de crecimiento se denomina fase exponencial, ya que se caracteriza por el crecimiento exponencial de las células. La velocidad de crecimiento durante esta fase se conoce como la tasa de crecimiento k y el tiempo que tarda cada célula en dividirse como el tiempo de generación g. Durante esta fase, los nutrientes son metabolizados a la máxima velocidad posible, hasta que dichos nutrientes se agoten, dando paso a la siguiente fase. La última fase de crecimiento se denomina fase estacionaria y se produce como consecuencia del agotamiento de los nutrientes en el medio. En esta fase las células reducen drásticamente su actividad metabólica y comienzan a utilizar como fuente energética aquellas proteínas celulares no esenciales. La fase estacionaria es un período de transición desde el rápido crecimiento a un estado de respuesta a estrés, en el cual se activa la expresión de genes involucrados en la reparación del ADN, en el metabolismo antioxidante y en el transporte de nutrientes.[108]
Genética Editar
Esquema de la conjugación bacteriana. 1-La célula donante genera un pilus. 2-El pilus se une a la célula receptora y ambas células se aproximan. 3-El plásmido móvil se desarma y una de las cadenas de ADN es transferida a la célula receptora. 4-Ambas células sintetizan la segunda cadena y regeneran un plásmido completo. Además, ambas células generan nuevos pili y son ahora viables como donantes.
La mayoría de las bacterias tienen un único cromosoma circular cuyo tamaño puede ir desde solo 160.000 pares de bases en la bacteria endosimbionte Candidatus Carsonella ruddii[109] a los 12.200.000 pares de bases de la bacteria del suelo Sorangium cellulosum.[110] Las espiroquetas del género Borrelia (que incluyen, por ejemplo, a Borrelia burgdorferi, la causa de la enfermedad de Lyme) son una notable excepción a esta regla pues contienen un cromosoma lineal.[111] Las bacterias pueden tener también plásmidos, pequeñas moléculas de ADN extra-cromosómico que pueden contener genes responsables de la resistencia a los antibióticos o factores de virulencia. Otro tipo de ADN bacteriano proviene de la integración de material genético procedente de bacteriófagos (los virus que infectan bacterias). Existen muchos tipos de bacteriófagos, algunos simplemente infectan y rompen las células huésped bacterianas, mientras que otros se insertan en el cromosoma bacteriano. De esta forma se pueden insertar genes del virus que contribuyan al fenotipo de la bacteria. Por ejemplo, en la evolución de Escherichia coli O157:H7 y Clostridium botulinum, los genes tóxicos aportados por un bacteriófago convirtieron a una inofensiva bacteria ancestral en un patógeno letal.[112] [113]
Imagen de un bacteriófago (virus que infecta bacterias).
Las bacterias, como organismos asexuales que son, heredan copias idénticas de genes, es decir, son clones. Sin embargo, pueden evolucionar por selección natural mediante cambios en el ADN debidos a mutaciones y a la recombinación genética. Las mutaciones provienen de errores durante la réplica del ADN o por exposición a agentes mutagénicos. Las tasas de mutación varían ampliamente entre las diversas especies de bacterias e incluso entre diferentes cepas de una misma especie de bacteria.[114] Los cambios genéticos pueden producirse al azar o ser seleccionados por estrés, en donde los genes implicados en algún proceso que limita el crecimiento tienen una mayor tasa de mutación.[115]
Las bacterias también pueden transferirse material genético entre células. Esto puede realizarse de tres formas principalmente. En primer lugar, las bacterias pueden recoger ADN exógeno del ambiente en un proceso denominado transformación. Los genes también se pueden transferir por un proceso de transducción mediante el cual un bacteriófago introduce ADN extraño en el cromosoma bacteriano. El tercer método de transferencia de genes es por conjugación bacteriana, en donde el ADN se transfiere a través del contacto directo (por medio de un pilus) entre células. Esta adquisición de genes de otras bacterias o del ambiente se denomina transferencia de genes horizontal y puede ser común en condiciones naturales[116] La transferencia de genes es especialmente importante en la resistencia a los antibióticos, pues permite una rápida diseminación de los genes responsables de dicha resistencia entre diferentes patógenos.[117]
Interacciones con otros organismos Editar
A pesar de su aparente simplicidad, las bacterias pueden formar asociaciones complejas con otros organismos. Estas asociaciones se pueden clasificar como parasitismo, mutualismo y comensalismo.
Comensales Editar
Debido a su pequeño tamaño, las bacterias comensales son ubicuas y crecen sobre animales y plantas exactamente igual a como crecerían sobre cualquier otra superficie. Así, por ejemplo, grandes poblaciones de estos organismos son las causantes del mal olor corporal y su crecimiento puede verse aumentado con el calor y el sudor.
Mutualistas Editar
Ciertas bacterias forman asociaciones íntimas con otros organismos, que les son imprescindibles para su supervivencia. Una de estas asociaciones mutualistas es la transferencia de hidrógeno entre especies. Se produce entre grupos de bacterias anaerobias que consumen ácidos orgánicos tales como ácido butírico o ácido propiónico y producen hidrógeno, y las arqueas metanógenas que consumen dicho hidrógeno.[118] Las bacterias en esta asociación no pueden consumir los ácidos orgánicos cuando el hidrógeno se acumula a su alrededor. Solamente la asociación íntima con las arqueas mantiene una concentración de hidrógeno lo bastante baja para permitir que las bacterias crezcan.
En el suelo, los microorganismos que habitan la rizosfera (la zona que incluye la superficie de la raíz y la tierra que se adhiere a ella) realizan la fijación de nitrógeno, convirtiendo el nitrógeno atmosférico (en estado gaseoso) en compuestos nitrogenados.[119] Esto proporciona a muchas plantas, que no pueden fijar el nitrógeno por sí mismas, una forma fácilmente absorbible de nitrógeno.
Muchas otras bacterias se encuentran como simbiontes en seres humanos y en otros organismos. Por ejemplo, en el tracto digestivo proliferan unas mil especies bacterianas. Sintetizan vitaminas tales como ácido fólico, vitamina K y biotina. También fermentan los carbohidratos complejos indigeribles y convierten las proteínas de la leche en ácido láctico (por ejemplo, Lactobacillus).[120] [121] [122] Además, la presencia de esta flora intestinal inhibe el crecimiento de bacterias potencialmente patógenas (generalmente por exclusión competitiva). Muchas veces estas bacterias beneficiosas se venden como suplementos dietéticos probióticos.[123]
Patógenos Editar
Micrografía electrónica con colores realzados que muestra a la especie Salmonella typhimurium (células rojas) invadiendo células humanas en cultivo.
Solo una pequeña fracción de las bacterias causan enfermedades en los seres humanos: de las 15.919 especies registradas en la base de datos de NCBI, solo 538 son patógenas.[124] Aún así son una de las principales causas de enfermedad y mortalidad humana, causando infecciones tales como el tétanos, la fiebre tifoidea, la difteria, la sífilis, el cólera, intoxicaciones alimentarias, la lepra y la tuberculosis. Hay casos en los que la etiología o causa de una enfermedad conocida se descubre solamente después de muchos años, como fue el caso de la úlcera péptica y Helicobacter pylori. Las enfermedades bacterianas son también importantes en la agricultura y en la ganadería, donde existen multitud de enfermedades como por ejemplo la mancha de la hoja, la plaga de fuego, la paratuberculosis, el añublo bacterial de la panícula, la mastitis, la salmonela y el carbunco.
Cada especie de patógeno tiene un espectro característico de interacciones con sus huéspedes humanos. Algunos organismos, tales como Staphylococcus o Streptococcus, pueden causar infecciones de la piel, pulmonía, meningitis e incluso sepsis, una respuesta inflamatoria sistémica que produce shock, vasodilatación masiva y muerte.[125] Sin embargo, estos organismos son también parte de la flora humana normal y se encuentran generalmente en la piel o en la nariz sin causar ninguna enfermedad.
Otros organismos causan invariablemente enfermedades en los seres humanos. Por ejemplo, el género Rickettsia, que son parásitos intracelulares obligados capaces de crecer y reproducirse solamente dentro de las células de otros organismos. Una especie de Rickettsia causa el tifus, mientras que otra ocasiona la fiebre de las Montañas Rocosas. Chlamydiae, otro filo de parásitos obligados intracelulares, contiene especies que causan neumonía, infecciones urinarias y pueden estar implicadas en enfermedades cardíacas coronarias.[126] Finalmente, ciertas especies tales como Pseudomonas aeruginosa, Burkholderia cenocepacia y Mycobacterium avium son patógenos oportunistas y causan enfermedades principalmente en las personas que sufren inmunosupresión o fibrosis quística.[127] [128]
Las infecciones bacterianas se pueden tratar con antibióticos, que se clasifican como bactericidas, si matan bacterias, o como bacterioestáticos, si solo detienen el crecimiento bacteriano. Existen muchos tipos de antibióticos y cada tipo inhibe un proceso que difiere en el patógeno con respecto al huésped. Ejemplos de antibióticos de toxicidad selectiva son el cloranfenicol y la puromicina, que inhiben el ribosoma bacteriano, pero no el ribosoma eucariota que es estructuralmente diferente.[129] Los antibióticos se utilizan para tratar enfermedades humanas y en la ganadería intensiva para promover el crecimiento animal. Esto último puede contribuir al rápido desarrollo de la resistencia antibiótica de las poblaciones bacterianas.[130] Las infecciones se pueden prevenir con medidas antisépticas tales como la esterilización de la piel antes de las inyecciones y con el cuidado apropiado de los catéteres. Los instrumentos quirúrgicos y dentales también son esterilizados para prevenir la contaminación e infección por bacterias. Los desinfectantes tales como la lejía se utilizan para matar bacterias u otros patógenos que se depositan sobre las superficies y así prevenir la contaminación y reducir el riesgo de infección.
La siguiente tabla muestra algunas enfermedades humanas producidas por bacterias:
Enfermedad Agente Principales síntomas
Brucelosis Brucella spp. Fiebre ondulante, adenopatía, endocarditis, neumonía.
Carbunco Bacillus anthracis Fiebre, pápula cutánea, septicemia.
Cólera Vibrio cholerae Diarrea, vómitos, deshidratación.
Difteria Corynebacterium diphtheriae Fiebre, amigdalitis, membrana en la garganta, lesiones en la piel.
Escarlatina Streptococcus pyogenes Fiebre, amigdalitis, eritema.
Erisipela Streptococcus spp. Fiebre, eritema, prurito, dolor.
Fiebre Q Coxiella burnetii Fiebre alta, cefalea intensa, mialgia, confusión, vómitos, diarrea.
Fiebre tifoidea Salmonella typhi, S. paratyphi Fiebre alta, bacteriemia, cefalalgia, estupor, tumefacción de la mucosa nasal, lengua tostada, úlceras en el paladar, hepatoesplenomegalia, diarrea, perforación intestinal.
Legionelosis Legionella pneumophila Fiebre, neumonía
Neumonía Streptococcus pneumoniae, Staphylococcus aureus,
Klebsiella pneumoniae, Mycoplasma spp., Chlamydia spp. Fiebre alta, expectoración amarillenta o sanguinolenta, dolor torácico.
Tuberculosis Mycobacterium tuberculosis Fiebre, cansancio, sudor nocturno, necrosis pulmonar.
Tétanos Clostridium tetani Fiebre, parálisis.
Clasificación e identificación Editar
Artículo principal: Clasificación científica
Cultivo de E. coli, donde cada punto es una colonia.
La clasificación taxonómica busca describir y diferenciar la amplia diversidad de especies bacterianas poniendo nombres y agrupando organismos según sus similitudes. Las bacterias pueden clasificarse con base en diferentes criterios, como estructura celular, metabolismo o con base en diferencias en determinados componentes como ADN, ácidos grasos, pigmentos, antígenos o quinonas.[131] Sin embargo, aunque estos criterios permitían la identificación y clasificación de cepas bacterianas, aún no quedaba claro si estas diferencias representaban variaciones entre especies diferentes o entre distintas cepas de la misma especie. Esta incertidumbre se debía a la ausencia de estructuras distintivas en la mayoría de las bacterias y a la existencia de la transferencia horizontal de genes entre especies diferentes,[132] la cual da lugar a que bacterias muy relacionadas puedan llegar a presentar morfologías y metabolismos muy diferentes. Por ello, y con el fin de superar esta incertidumbre, la clasificación bacteriana actual se centra en el uso de técnicas moleculares modernas (filogenia molecular), tales como la determinación del contenido de guanina/citosina, la hibridación genoma-genoma o la secuenciación de ADN ribosómico, el cual no se ve involucrado en la transferencia horizontal.[133]
El Comité Internacional de Sistemática de Procariotas (ICSP) es el organismo encargado de la nomenclatura, taxonomía y las normas según las cuales son designados los procariotas.[134] El ICSP es responsable de la publicación del Código Internacional de Nomenclatura de Bacterias (lista de nombres aprobados de especies y taxones bacterianos).[135] También publica la Revista Internacional de Bacteriología Sistemática (International Journal of Systematic Bacteriology).[136] En contraste con la nomenclatura procariótica, no hay una clasificación oficial de los procariotas porque la taxonomía sigue siendo una cuestión de criterio científico. La clasificación más aceptada es la elaborada por la oficina editorial del Manual de Bacteriología Sistemática de Bergey (Bergey's Manual of Systematic Bacteriology) como paso preliminar para organizar el contenido de la publicación.[137] Esta clasificación, conocida como "The Taxonomic Outline of Bacteria and Archaea" (TOBA), está disponible en Internet.[138] Debido a la reciente introducción de la filogenia molecular y del análisis de las secuencias de genomas, la clasificación bacteriana actual es un campo en continuo cambio y plena expansión.[139] [140]
La identificación de bacterias en el laboratorio es particularmente relevante en medicina, donde la determinación de la especie causante de una infección es crucial a la hora de aplicar un correcto tratamiento. Por ello, la necesidad de identificar a los patógenos humanos ha dado lugar a un potente desarrollo de técnicas para la identificación de bacterias.
Streptococcus mutans visualizado con la tinción de Gram. Cada pequeño punto de la cadena es una bacteria.
La técnica de tinción de membranas de bacterias de Gram, desarrollada por Hans Christian Gram en 1884,[141] ha supuesto un antes y un después en el campo de la medicina, y consiste en teñir con tintes específicos diversas muestras de bacterias en un portaobjetos para saber si se han teñido o no con dicho tinte.[142]
Una vez se han adicionado los tintes específicos en las muestras, y se ha lavado la muestra pasados unos minutos para evitar confusiones, hay que limpiarlas con unas gotas de alcohol etílico. La función del alcohol es la de eliminar el tinte de las bacterias, y es aquí donde se reconocen las bacterias que se han tomado: si la bacteria conserva el tinte, es una Gram positiva, las cuales poseen una pared más gruesa constituida por varias decenas de capas de diversos componentes proteicos; en el caso de que el tinte no se mantenga, la bacteria es una Gram negativa, la cual posee una pared de una composición diferente. La función biológica que posee ésta técnica es la de fabricar antibióticos específicos para esas bacterias.
Esta tinción es empleada en microbiología para la visualización de bacterias en muestras clínicas. También se emplea como primer paso en la distinción de diferentes especies de bacterias,[143] considerándose bacterias Gram positivas a aquellas que se tornan de color violeta y Gram negativas a las que se tornan de color rojo.[144] [145]
En el análisis de muestras clínicas suele ser un estudio fundamental por cumplir varias funciones:
Identificación preliminar de la bacteria causante de la infección.
Consideración de la calidad de la muestra biológica para el estudio, es decir, permite apreciar el número de células inflamatorias así como de células epiteliales. A mayor número de células inflamatorias en cada campo del microscopio, más probabilidad de que la flora que crezca en los medios de cultivo sea la representativa de la zona infectada. A mayor número de células epiteliales sucede los contrario, mayor probabilidad de contaminación con flora saprófita.
Utilidad como control de calidad del aislamiento bacteriano. Las cepas bacterianas identificadas en la tinción de Gram se deben corresponder con aislamientos bacterianos realizados en los cultivos. Si se observan mayor número de formas bacterianas que las aisladas, entonces hay que reconsiderar los medios de cultivos empleados así como la atmósfera de incubación.
Filos y filogenia Editar
Probable modelo evolutivo de los principales filos y clados.
Artículo principal: Filogenia bacteriana
Las relaciones filogenéticas de los seres vivos son motivo de controversia y no hay un acuerdo general entre los diferentes autores. La mayoría de árboles filogenéticos, en especial los de ARNr 16S y 23S, muestran que los grupos basales son filos termófilos como Aquificae y Thermotogae,[146] lo que reforzaría el origen termófilo de los dominios Archaea y Bacteria. En cambio, algunos árboles genómicos muestran a Firmicutes (Gram positivos) como el clado más antiguo.[147] Según las teorías de Cavalier-Smith la mayor divergencia se encuentra en un grupo fotosintético que denomina Chlorobacteria (Chloroflexi).[37] Otros estudios filogenéticos genómicos o proteicos colocan en una posición basal a Planctomycetes, Proteobacteria u otros filos. Finalmente se ha propuesto que hubo una temprana divergencia entre dos supergrupos: Gracilicutes y Terrabacteria;[148] demostrando en suma que actualmente no existe un filogenia bacteriana estable como para conocer con certeza la historia evolutiva bacteriana más temprana. Esto debido con toda probabilidad al fenómeno de la transferencia genética horizontal, típica de los organismos procariotas.
Los principales filos bacterianos se pueden organizar dentro de un amplio criterio filogenético en tres conjuntos:
Grupos termófilos Editar
Venenivibrio, una bacteria aquifical termófila de aguas termales.
De acuerdo con la mayoría de árboles filogenéticos moleculares, los filos termófilos son los más divergentes, formando un grupo parafilético basal, lo que es compatible con las principales teorías sobre el origen y evolución procariota. Son termófilos e hipertermófilos con metabolismo quimiotrofo, respiración anaerobia y estructura Gram negativa (de doble membrana), destacando los siguientes filos:
Aquificae. Pequeño grupo de bacterias quimiolitotrofas, termófilas o hipertermófilas. Se las encuentra en manantiales calientes, pozos sulfurosos y fuentes hidrotermales oceánicas.
Thermotogae. Un filo de hipertermófilos, anaerobios obligados, heterótrofos fermentativos.
Dictyoglomi. Comprende una sola especie de hipertermófilo, quimioorganotrofo y aerobio.
Thermodesulfobacteria: Termófilas reductoras de sulfato.
Caldiserica. Bacteria termófila anaerobia.
Synergistetes. Bacterias anaerobias. Aunque pocos géneros son termófilos, Synergistetes tiene posición basal en la filogenia bacteriana del ARNr.
Gram positivos y relacionados Editar
Tinción de Gram de Bacillus anthracis, una bacteria patógena Gram positiva del filo firmicutes.
Los grupos Gram positivos son básicamente Firmicutes y Actinobacteria, los cuales habría engrosado su pared celular como una adaptación a la desecación con pérdida de la membrana externa, desarrollando esteroles, ácido teicoico y formando esporas en varios grupos. El término Posibacteria se ha usado como taxón para agrupar a los Gram positivos y grupos derivados como Tenericutes. El término monodérmico alude a la única membrana celular que poseen los Gram positivos, lo que significa que otros filos como Chloroflexi y Thermomicrobia, al ser monodérmicos, están relacionados con los primeros a pesar de que son Gram variables. Según algunos árboles filogenéticos, los filos monodérmicos forman parte de un superclado denominado Terrabacteria, llamados así por su probable evolución en medios terrestres, y se incluye en él a filos didérmicos como Deinococcus-Thermus que es Gram variable y Cyanobacteria que es Gram negativo. Los Gram positivos y relacionados (Terrabacteria) se presentan en la mayoría de árboles filogenéticos como un grupo parafilético con respecto a Gracilicutes y está conformado por los siguientes filos:
Actinobacteria. Un extenso filo de bacterias Gram positivas de contenido GC alto. Son comunes en el suelo aunque algunas habitan en plantas y animales, incluyendo algunos patógenos. Algunas forman colonias en forma de hifas (Actinomyces).
Firmicutes o Endobacteria. Es el grupo más extenso y comprende a las bacterias Gram positivas con contenido GC bajo. Se encuentran en diversos hábitats, incluyendo algunos patógenos notables. Una de las familias, Heliobacteria, obtiene su energía a través de la fotosíntesis y otros tienen una pseudo membrana externa (Negativicutes).
Tenericutes o Mollicutes. Son endosimbiontes Gram negativos monodérmicos y sin pared celular. Se derivan de Firmicutes según la mayoría de filogenias.
Deinococcus-Thermus o Hadobacteria. Pequeño grupo de quimiorganotrofos extremófilos altamente resistentes. Unas especies soportan el calor y el frío extremo, mientras que otras son resistentes a la radiación y a las sustancias tóxicas.
Chloroflexi. Pequeño filo de bacterias monodérmicas Gram variables aerobias facultativas y típicamente filamentosas. Incluye a las bacterias verdes no del azufre, las cuales realizan la fotosíntesis anoxigénica mediante bacterioclorofila (sin producción de oxígeno) y en donde su vía de fijación del carbono también difiere de la de otras bacterias fotosintéticas.
Thermomicrobia. Pequeño filo (o clase) termófilo derivado de Chloroflexi.
Cyanobacteria (algas verde-azuladas). El grupo más importante de bacterias fotosintéticas. Presentan clorofila y realizan la fotosíntesis oxigénica. Son unicelulares o coloniales filamentosas.
Nitrospirae. Grupo de quimiosintéticos oxidantes de nitrógeno y algunos son termófilos.
En algunas filogenias aisladas se relaciona también a Fusobacteria, Armatimonadetes y Dictyoglomi.
Gracilicutes Editar
Las espiroquetas, al igual que otros Gracilicutes, son Gram negativas.
El superclado Gracilicutes o Hydrobacteria está bien consensuado en muchos árboles filogenéticos. Son el mayor grupo de bacterias Gram negativas, didérmicas, en su mayoría quimioheterótrofas, de hábitat acuático o relacionado con animales y plantas como comensal, mutualista o patógeno. Está conformado por varios filos y superfilos:
Spirochaetes. Bacterias quimioheterótrofas con forma alargada típicamente enrollada en espiral que se desplazan mediante rotación. Muchas producen enfermedades.
Grupo FCB o Sphingobacteria.
Fibrobacteres. Pequeño filo de que incluye muchas de las bacterias estomacales que permiten la degradación de la celulosa en los rumiantes.
Chlorobi. Destacan las bacterias verdes del azufre, las cuales son fototrofas mediante bacterioclorofila y anaerobias obligadas. Algunas son termófilas que viven en fuentes hidrotermales.
Bacteroidetes. Un extenso filo de bacterias con amplia distribución en el medio ambiente, incluyendo el suelo, sedimentos, agua de mar y el tracto digestivo de los animales. Es un grupo heterogéneo que incluye aerobios obligados o anaerobios obligados, comensales, parásitos y formas de vida libre.
Gemmatimonadetes. Aerobios del suelo y el fango.
Grupo PVC o Planctobacteria
Planctomycetes. Bacterias principalmente acuáticas aerobias encontradas en agua dulce, salobre y marina. Su ciclo biológico implica la alternancia entre células sésiles y flageladas. Se reproducen por gemación.
Verrucomicrobia. Comprende bacterias terrestres, acuáticas y algunas asociadas con huéspedes eucariotas.
Chlamydiae. Un pequeño grupo de parásitos intracelulares obligados de las células eucariotas.
Lentisphaerae. Pequeño grupo de bacterias recientemente descubiertas en aguas marinas y hábitats terrestres anaerobios.
Proteobacteria (bacterias púrpuras y relacionadas). Es un grupo muy diverso y extenso. La mayoría son heterótrofas, otras son fermentadoras como las enterobacterias y muchas causan enfermedades como las ricketsias que son parásitos intracelulares. Los rizobios son endosimbiontes fijadores de nitrógeno en las plantas, las bacterias púrpuras son fototrofas con bacterioclorofila y las mixobacterias forman agregados multicelulares. Algunos autores consideran que son derivados o relacionados con Proteobacteria los siguientes filos:
Acidobacteria. Pequeño filo de bacterias acidófilas comunes en el suelo. Incluye una bacteria fototrofa usando bacterioclorofila.
Armatimonadetes. Pequeño grupo aerobio quimioheterótrofo.
Elusimicrobia. Se encuentra disperso por mar, tierra y como endosimbionte de insectos.
Deferribacteres. Pequeño grupo de bacterias acuáticas anaerobias.
Chrysiogenetes. Pequeño grupo quimiolitoautótrofo anaerobio con una bioquímica y una forma de vida únicas, capaces de respirar arseniato.
Fusobacteria. No siempre se le incluye en Gracilicutes. Son bacterias heterótrofas anaerobias causantes de infecciones en humanos. Constituyen uno de los principales tipos de flora del aparato digestivo.
Uso de las bacterias en la tecnología y la industria Editar
Muchas industrias dependen en parte o enteramente de la acción bacteriana. Gran cantidad de sustancias químicas importantes como alcohol etílico, ácido acético, alcohol butílico y acetona son producidas por bacterias específicas. También se emplean bacterias para el curado de tabaco, el curtido de cueros, caucho, algodón, etc. Las bacterias (a menudo Lactobacillus) junto con levaduras y mohos, se han utilizado durante miles de años para la preparación de alimentos fermentados tales como queso, mantequilla, encurtidos, salsa de soja, chucrut, vinagre, vino y yogur.[149] [150]
Las bacterias tienen una capacidad notable para degradar una gran variedad de compuestos orgánicos, por lo que se utilizan en el reciclado de basura y en biorremediación. Las bacterias capaces de degradar los hidrocarburos son de uso frecuente en la limpieza de los vertidos de petróleo.[151] Así por ejemplo, después del vertido del petrolero Exxon Valdez en 1989, en algunas playas de Alaska se usaron fertilizantes con objeto de promover el crecimiento de estas bacterias naturales. Estos esfuerzos fueron eficaces en las playas en las que la capa de petróleo no era demasiado espesa. Las bacterias también se utilizan para la biorremediación de basuras tóxicas industriales.[152] En la industria química, las bacterias son utilizadas en la síntesis de productos químicos enantioméricamente puros para uso farmacéutico o agroquímico.[153]
Las bacterias también pueden ser utilizadas para el control biológico de parásitos en sustitución de los pesticidas. Esto implica comúnmente a la especie Bacillus thuringiensis (también llamado BT), una bacteria de suelo Gram-positiva. Las subespecies de esta bacteria se utilizan como insecticidas específicos para lepidópteros.[154] Debido a su especificidad, estos pesticidas se consideran respetuosos con el medio ambiente, con poco o ningún efecto sobre los seres humanos, la fauna y la mayoría de los insectos beneficiosos, como por ejemplo, los polinizadores.[155] [156]
Cristales de insulina.
Las bacterias son herramientas básicas en los campos de la biología, la genética y la bioquímica moleculares debido a su capacidad para crecer rápidamente y a la facilidad relativa con la que pueden ser manipuladas. Realizando modificaciones en el ADN bacteriano y examinando los fenotipos que resultan, los científicos pueden determinar la función de genes, enzimas y rutas metabólicas, pudiendo trasladar posteriormente estos conocimientos a organismos más complejos.[157] La comprensión de la bioquímica celular, que requiere cantidades enormes de datos relacionados con la cinética enzimática y la expresión de genes, permitirá realizar modelos matemáticos de organismos enteros. Esto es factible en algunas bacterias bien estudiadas. Por ejemplo, actualmente está siendo desarrollado y probado el modelo del metabolismo de Escherichia coli.[158] [159] Esta comprensión del metabolismo y la genética bacteriana permite a la biotecnología la modificación de las bacterias para que produzcan diversas proteínas terapéuticas, tales como insulina, factores de crecimiento y anticuerpos.[160] [161]
Comentarios (7)

- 1. | 15/06/2025

- 2. | 14/06/2025

- 3. | 08/06/2025

- 4. | 17/04/2025

- 5. | 05/04/2025

- 6. | 21/03/2025
one of the major purposes of the train is to maximise
the pump. Yes, the movement will nonetheless build mass, but
you won’t have the power to raise as heavy as you
can on regular curls. That’s why the train options prominently
within the full bicep exercises beneath. Reverse curls are top-of-the-line
workouts for biceps and brachioradialis improvement that you are in a place to do within the fitness
center.
Coaching all three muscular tissues ensures a balanced arm, aesthetically and functionally, enhancing
your overall strength and definition. Superset the pairs of workouts famous,
resting solely after you complete each moves. Now the simplest
method to discover out which head is lagging is literally just flex like within the mirror, and see which aspect appears
less developed. Or, just check out both workout routines
and see which one you find more difficult and provides you the better
pump. You must position your arm so that the effort you’re placing is directed consistent with the realm you’re trying to focus on. And the short head runs down the inside — providing you with more of that inner
peak. Count On expert-backed exercises, vitamin advice, the newest in energy sports, and a complete lot of motivation heading your means.
Perform the 21s on the end of your workout for a muscle-ripping pump.
Use a lightweight, as you’ll be fatigued,
to ensure you follow the proper form. Carry Out the lying cable curl on the finish of your workout
for a muscle-ripping pump. Chin-ups are an excellent compound (multi-joint) train usually accomplished at the beginning of an upper-body
workout. You might also do them as the second exercise in a superset to smoke your guns.
The cable crossover curl exercise mimics a double overhead biceps pose — just
like a bodybuilder.
Second, given the significance of the stretch, your arm
must be almost fully extended on the backside. However that is usually a
place that most people have by no means skilled
their biceps in. So the factor about a standing dumbbell curl is it’s hardest
within the center right around here. There are only
four workout routines, however proper kind is crucial, so pay attention if
you need to grow your arms. As mentioned, most individuals store
comparatively little physique fat on their arms, so even bringing your
total physique fats down by a few p.c can make your
biceps look learner and extra outlined. So generally, you might actually
have properly defined biceps already; it’s simply that you might not notice this definition because
part of your biceps is underdeveloped.
After making an attempt over one hundred fifty workout routines and testing numerous pairings, we got here up with a listing of the 7
greatest bicep supersets for hypertrophy. So if you wish to get a great bicep pump and add mass to your arms, then these supersets will do
the trick after which some. The cable curl carefully resembles the barbell curl, with
the distinction that you might find it simpler to
keep fixed pressure in your biceps with this exercise.
Your biceps are one of the visually striking muscle tissue, and has been an icon of bodybuilding for decades.
Few muscle tissue garner more coaching attention than these, and
given how they're all the time seen if you end up in brief sleeves, that is comprehensible.
But for larger achieve, It is crucial to extend the stress
positioned on the muscles progressively and train numerous muscle fibers with different workout routines.
Barbell workout routines are effective at recruiting your arm, which can be used to
build measurement, as long as they're accomplished at an appropriate frequency and volume.
The brachialis muscle is among the major muscles
within the upper arm.
Even inside the completely different workout routines, mess around with totally
different grips. In order to train the biceps completely, you need to hit the brachialis and the brachioradialis,
which means you must use a pronated and neutral grip in your training.
In addition, you can also fiddle with impartial and extensive grips.
As mentioned above, embody other bicep-oriented workout routines different
than just arm curls. Choose huge compound lifts that will hit the biceps to a larger diploma.
However with a bar, you actually want to use close grip, which will
flare your arms inwards. The solely potential draw back with this exercise is that it’s most challenging close to the highest place of the curl when the biceps are
nearly totally contracted. However should you don’t have access to cables, you can always replicate this motion with dumbbells.
Muscle activation isn’t the be-all and end-all for muscle growth.
It Is nice this target the lengthy head, however
ensure different variations you include are extra focussed on the lateral and medial head.
Sure, barbells may help you build arm muscle tissue with each rep, speeding up and simplifying your
arm workouts. The barbell incline tricep extension helps to construct
muscle and strength in all three tricep heads. The shut grip bench press is another nice exercise you can add to your barbell arm workout routines.
The mendacity barbell tricep extension is one other great exercise for building strength in your arms.
Right Here are 12 of the most effective barbell bicep and tricep workout routines to construct muscle mass.
So listed beneath are the four workouts you should embody in your arm exercise to utterly blow up your arms — complete with the reps,
sets, and alternatives.
Now it’s time to pick up the iron and use
that data to construct the big and strong biceps you’ve always needed.
This workout combines barbell, dumbbell, and cable workout
routines into a balanced mix. The heavier workouts early
in the exercise lend themselves to decrease reps, while finishing off with a couple of high-rep units ends the workout on a excessive observe and offer you a sleeve-bursting pump.
The cable curl is similar to the barbell curl in execution and movement but with a bar hooked up to a
cable machine. Make sure you’re not bringing your shoulder forward as you curl the weights up.
So not only are you going to discover methods to get more outlined biceps via dietary and body-fat-reduction means; you’ll find out about the best bicep workouts for the job as
well. The greatest approach to do a 3-day full physique exercise is to alternate days and alternate
workouts. You ought to cycle through a minimum of two
different full body workouts and do them on alternating days.
For instance, on Monday perform Workout A, on Wednesday carry
out Workout B and Friday Perform Exercise A. Take two days
of relaxation and begin off the following Monday with exercise B.
Full physique workouts are an efficient way to construct muscle as a outcome of they have an inclination to concentrate on massive heavy lifts that
provide you with an excellent opportunity for progressive
overload. Instead of thinking about which precise lift goes to hit which main muscle groups,
I need you to suppose more when it comes to movement sorts.
If we will train every of the most important compound movements in one efficient exercise, then we’ll be incorporating the muscles that
achieve those movement patterns.
This seated variation uses dumbbells, so you’ll need to work additional onerous
to carry and stabilize the weights. This train will produce a deep burn and pump in all three deltoid heads.
When performing any of those exercises, keep in mind to focus on yourself and never different lifters.
Do an adequate heat up, take your time getting the right
starting place and perform the workout routines with correct type via the complete vary of motion. As Soon As you've
been performing again and biceps exercises or different power coaching workouts
for a minimum of six months you should have little question been able to build muscle.
A bulging pair of bis is a logo of strength that grabs people’s attention and enhances your physique.
By applying these criteria, Nippard was able to separate the most effective muscle-building workouts from
the ineffective ones—leading to his ultimate
rankings. One of the best biceps moves for hitting
the brief head of the biceps is the No Cash Curl.
We wish to ensure we are adequately hitting the beginning portion off the biceps’
vary of motion.
These exercises add more risk to damage due to the excessive volume of 7
sets. [newline]These barbell arm workouts are extremely
recommended for anybody interested in building arm muscles and gaining power.
Barbell spider curls practice your biceps in a shortened (which
is to say, a maximally contracted) muscle position. This leads to a robust muscle
pump that may make your arms appear more vascular and result
in vital hypertrophy. If you’re looking for simple barbell arm workouts to add to your routine, barbell curls are
a fantastic staple train to get you began. If you need to construct bigger arms using a barbell, you must work out your
biceps, triceps, and forearms. That is why we now have selected workout routines
that hit all of those muscle teams. If your biceps look good
when flexed but a bit skinny when they’re relaxed, then you
can do a selected lengthy head bicep workout to beef up the outer muscle fibers of your
bis.
If you need a big biceps peak, this is doubtless certainly one of the best biceps
exercise. To construct muscular tissues within the
upper arms, cable curls must be persistently applied into energy
training arm workouts. It's good to do a full physique power exercise 3 times per week for the goal of building
muscle as a end result of it provides enough relaxation time
between coaching sessions in your muscles to recuperate and develop.
Three instances per week is a perfect frequency for muscle growth if you are doing complete
physique workouts. It Is best to do full body exercise routines about thrice a week
as a result of it allows for adequate rest intervals between workouts for the aim
of constructing muscle. Three days per week is the perfect
frequency of exercises for muscle progress as a outcome of you could have adequate rest time between coaching to
allow for the muscle constructing response.
Who doesn’t need biceps that look great in a tank prime and can do
spectacular things (like wield Thor’s hammer or Captain America’s shield)?
Doing regular body weight workouts will strengthen your biceps — and it
can’t hurt your probabilities of changing into the subsequent Avenger.
Doing supersets is a wonderful way to save time and get an even higher muscle pump.
A chest and bicep superset session works higher than supersetting chest and triceps because the muscle groups don’t intervene with one another.
The preacher curl ought to be your go-to train when you wish to isolate your biceps
fully. You place your higher arm in a set place, making it inconceivable
to cheat or to make use of other muscle tissue to help curl the load up.
The biceps brachii (or simply the biceps for short) is a two-headed muscle situated at the front of the upper
arm. The two heads (the quick and the long head) start at different spots on your shoulder blade and be a part of together on the elbow.
Whereas we went over the compound workouts, keep in thoughts that you continue to need to use
this same idea to the biceps.
Here, I’ll break down the 10 greatest barbell biceps curls, explaining how every one targets your
arms from a unique angle for enhanced biceps development. Most analysis reveals
you'll find a way to maximize your price of muscle growth with about 10–20 units per week.
The higher limit isn’t very nicely established but, especially when you’re
attempting to focus on only a few muscle teams at a time.
It’s attainable that you could grow your biceps even faster by doing 30
and even 40 sets per week, especially when you work as a lot
as it steadily. To train both heads, I advocate beginning together with your favourite vertical
pull. Chin-ups/pulldowns with an underhand, angled, or impartial grip give your biceps higher
leverage, and they could also be higher.
Rest 2–3 minutes before shifting on to the dumbbell incline press.
You can obtain a PDF file of all of the workouts and the routine if you
would like. The susceptible lying position also helps
hold the momentum in management and allows you to have interaction the
again muscles effectively.
She is presently studying to turn into a private trainer and
hopes to begin teaching in 2022. Chin-ups are some of the difficult
and effective workout routines you are capable of do.
Do one exercise after the other with seconds in-between workouts.
As such, you'll be working all your main muscle teams, whether isotonically or isometrically.
Start light and use hex or 12-sided sort of dumbbells because the spherical ones will only
make things harder to stabilize. Steadily increase the weight as you get stronger to continue
progressing.
The plank hits your glutes, core, chest, triceps and shoulders
in an isometric trend. The renegade row (which is a row from
a plank position) works your biceps, again, and rear delts.
And the plank to squat works your legs, glutes, low again and
traps. It’s a great complex, multi-phase, multi-joint exercise that's going that can help you
construct mass and brute power whereas burning a lot of calories at the identical time.
The clean is also an train that can be done with comparatively heavy dumbbells.
The #1 train for training biceps is the curl, and there are several curl variations that emphasize
totally different bicep muscle functions and parts of the power
curve. You can incorporate many different variations
of the curl right into a beginner bicep exercise, or when you want
a complicated bicep workout you ought to use techniques that increase the
level of depth. For your dumbbell biceps exercise, begin with a moderate weight and you may add further weight over time as
you build energy and start to raise at an advanced level.
We all know guys who can do tons of pull ups and heavy rows, however nonetheless don’t have the best biceps
muscle mass. Some bicep curl machines allow you to change grips,
meaning you could also do machine reverse or hammer curls. Regardless of construction, the machine bicep curl is a
safe and efficient isolation train for your higher arms.
This limits its effectiveness considerably in fully creating the biceps, significantly in the elongated vary of
movement, however it stays a staple for constructing mass and
foundational power. They are generally performed after the barbell curl; nevertheless, you'll find a way
to carry out them at any point throughout your exercise.
You should carry out the barbell biceps curl firstly of your
arm coaching routine. Nevertheless, should you train your back and biceps on the same day, you must do it after you full your again routine.
You could know some of them, but some of these exercises are
distinctive.
For most reps, be certain to use correct kind and don’t swing your arms or
body to get the bar going. However, be at liberty to make
use of slightly momentum on the last repetition of your units.
Doing so lets you get previous the sticking level and do
one more rep than you'd have in any other case. Used sparingly, the
"cheat curl" is an efficient way to overload your biceps and
force them to develop. The cable reverse curl, also known as the reverse cable curl, is an exercise that targets the brachioradialis muscle
within the forearms, along with the biceps and different forearm muscles.

- 7. | 19/03/2025
constant foundation compared to having to devour these calories from whole
foods,’ Fleming provides. Now as for what quantity of drop sets to do to
build muscle quick, one study replaced three regular sets with 1 normal set followed by 2 drop units.
However for now, this is an instance of what this could seem like with a 3-day per
week full body workout that might train each muscle with around 6 units per week.
Now, the final research I want to take a glance at
earlier than providing some suggestions for the least
variety of sets you are capable of do to build muscle quick is a meta-analysis led by Brad Schoenfeld.
The first technique that'll allow you to construct muscle quick in much less time has
to do together with your exercise quantity. It is recommended that you simply give consideration to slowly increasing your weights to not more than 10%
every week so the physique can adapt. This also can help scale back your threat of damage or overtraining.
That is something you can’t guarantee with different brands cut-rate, over-processed blends.
In Accordance to a 2021 review, females have 70% to 80% decrease stores of creatine than males.
Researchers additionally observe that females metabolize creatine in one other way.
Creatine stores could drop during hormone-related changes, similar to menstruation, being
pregnant, and menopause. The traits of creatine
differ between women and men. If you train often, you likely need to ensure you’re
getting the most out of it. Genetics play an essential role in figuring out your body's capacity to
put on muscle (and its limitations), partly by influencing your hormonal and muscular make-up.
These small rubber attachments increase the diameter of a dumbbell or barbell deal with, growing muscular contractions.
These can help customers become stronger and acquire extra mass in upper-body
muscle groups as a outcome of overloading the muscular tissues with a model
new adaptive stimulus. In our experience, coaching with
Fat Gripz is among the easiest ways to overload the
muscles and add new muscle tissue. If people aren’t gaining vital measurement on their chest, arms, and thighs but are gaining plenty around their waist, regulate calorie consumption by consuming in a smaller surplus.
We discover that ‘hard gainers’ typically profit from including cheat meals to their diet
to increase their general calorie intake.
It is tough to eat four,000 calories worth of chicken and rice, but
this turns into significantly simpler with the addition of a pizza or a
burger and fries.
You store key nutrients in your muscular tissues,
together with glycogen (primarily from carbs). And having more
muscle often means you can course of more dietary carbs efficiently and
utilize your calories higher. Attain your full muscle-building potential with this free
meal prep toolkit. Save time and money in your meal prep, and maximize features
with this easy step-by-step information.
Additionally, when first starting out, you may not see main features in measurement immediately.
To fix this, attempt taking a fiber complement at evening, proper before going to bed.
This way, it won’t intervene together with your caloric consumption targets.
Optimum Diet practically places a multivitamin into each serving of Severe Mass; your every day requirements for many vitamins and minerals are met or exceeded in a single 1250 calorie serving.
HMB is answerable for a number of the useful effects
of protein and leucine within the diet. It could also be especially important for
decreasing the breakdown of muscle proteins.
Nevertheless, your training ought to include compound and
isolation movementsfor one of the best long-term fitness outcomes.
You should not often have more than "two reps within the tank" by the end of a set in case
your objective is constructing muscle. To put it simply, relying on your size,
your muscle tissue might grow extra with lower reps using heavy weights or with high reps
utilizing lighter weights.
Although you will mechanically practice surrounding muscle
tissue and this approach is not probably the
most environment friendly or beneficial approach to add mass.
Based Mostly on existing science and evidence-based practices, listed
right here are the necessary thing parts you
must be specializing in in terms of curating your muscle constructing game plan.
Soreness can last as lengthy as a number of days after coaching but is not essentially a cause to stop
training altogether. Some relaxation and recovery are needed, however coaching on sore muscles won't cause any extra hurt - in some circumstances,
continued training can help relieve muscle pain, particularly utilizing
lively recovery.
Full-body exercises allow your body sufficient
restoration as a end result of it’s beneficial you take no less than one day off between workouts to allow your physique to completely relaxation and recover.
With split routines, you must work out a quantity of days in a row, or
you won’t be in a position to work all of your muscle tissue each week.
Though slicing out carbohydrates might help you shed weight
temporarily, your testosterone, energy, and muscle growth takes a toll (31).
It’s crucial that when you're trying to construct muscle, carbs are a major source of your whole every day calories.
Since compound lifts work bigger muscular tissues,
you should rest for a longer time. When you practice your accent muscular tissues,
like triceps and biceps, take shorter relaxation durations to maximise hypertrophy.
How you structure your coaching, your alternative of workouts, your food plan, your recovery strategies, and your mental perspective all play important
roles in your quest to pack on the lean muscle mass.
Periodization is a coaching precept that places you through totally different training phases.
For instance, phase I might concentrate on power, section II might focus on power coaching, and part
III could focus on hypertrophy. Every part is usually several weeks lengthy and allows for a wide range of
movements, reps, units, and workout protocols so that your
physique by no means adapts and retains growing.
Pump coaching is defined as high-volume coaching which causes sarcoplasmic hypertrophy, a rise in sarcoplasmic
fluid in your cells. To construct muscle, you need to
place enough stress in your muscles to force them to adapt; and lifting weights is the right
answer.
You could additionally be getting stronger but not growing larger muscular tissues if you’re specializing in strength
coaching rather than hypertrophy training. This means
you might be lifting heavier weights for 1 to five reps as a substitute of lighter weights for six to 12 reps.
Your rests may also be longer between units. The different two main macronutrients are, in fact, carbohydrates and protein. By
offering an enormous quantity of calories, mass gainer powder could be simply combined into water to successfully
add hundreds or 1000's of energy per day into your diet.
Excessive quality mass gainers often supplement the calories with
further vitamins and minerals so that you aren’t dealing with any
deficiencies.
If you wish to bulk up a certain muscle group, train it extra typically to increase muscle
nuclei contained in the cell. Overtraining is actual; nonetheless, it applies to the
central nervous system (not specifically to muscles) and is essentially
exaggerated in bodybuilding. If individuals have issues digesting milk and prefer to consume whey protein powder
in water, they can add 40 grams of maltodextrin or dextrose to a
post-workout shake. We discover this provides rapid replenishment of glycogen stores.
In Accordance to analysis, resistance training locations your muscles underneath sufficient rigidity and plays a vital position in muscle development (1).
Mass Gainers do not comprise any harmful ingredients,
so there's nothing to fret about there. As properly
as various bodily circumference measurements like your arms, chest, shoulders, and legs, for
instance. Ideally, your energy and these bodily measurements ought to be gradually growing alongside together with
your physique weight.
A small 2020 research with postmenopausal women found that 8
weeks of resistance training improved muscle mass and strength.
Nonetheless, including BCAA dietary supplements did not obtain any significant gains.
A Quantity Of dietary supplements can help assist muscle progress when paired with resistance coaching and a well-rounded diet.
This consists of creatine and protein dietary supplements, amongst others.
Unleash your physique's untapped growth potential with Anafuse, the breakthrough pure anabolic that delivers seen muscle size and
strength features without undesirable side effects. Customers constantly
report breaking via plateaus inside weeks, including
quality mass that doesn't disappear when cycling off, making this the proper introduction to critical mass-building supplementation.
Clear Labs Mass Gainer does the best job of any mass gainer in the
marketplace, supplying three grams of creatine per serving.
To achieve muscle, you should eat enough calories and protein, as
well as train, ideally with weights. Once your vitamin and exercise regimens are in verify, you might wish to contemplate dietary dietary supplements.
A 2009 research reports that including a beta-alanine
supplement to a 6-week, high intensity interval coaching program increased lean body mass by about 1
lb (0.45 kg) more than a placebo in male participants.
Weight gainers are dietary supplements designed to conveniently help you get
more calories and protein. They’re sometimes utilized by people who
have issue gaining muscle, even when consuming giant amounts of
calories and lifting weights.
In addition, protein offers less than half as many calories per gram as fat
- fats supplies nine energy per gram, compared to protein which supplies only
four energy per gram. Slow-twitch are used to assist endurance
training like jogging or cycling, and fast-twitch
are used in extra explosive and fast burst exercise (21,22).
Fast-twitch fibers usually have a tendency to enhance in measurement in comparability with slow-twitch fibers as a
outcome of fast-twitch fibers fatigue extra shortly, resulting in an increased want for
repair and strengthening.
Not that you simply wish to rush through your workouts,
however you want to have the identical depth
stage as a sprinter. Your mindset on this workout schedule should be to
get in, blast your muscle tissue like hell after which
get out. If you have a set goal of someone’s physique you would like to emulate, put a poster of
them in your wall, set your phone background to them, or before
you prepare, watch videos of them lifting weights.
By following these guidelines, you'll have the
ability to achieve your muscle growth objectives
and revel in a more healthy, stronger physique. Stay dedicated, monitor your progress, and modify as wanted to see the best outcomes.
Consistency is vital to achieving desired muscle development with mass gainers.
Monitoring weight and body composition helps in adjusting the dosage of mass gainers effectively.
Guarantee protected and efficient use of mass
gainers by following really helpful dosages to avoid extreme fats acquire and
mixing them with a balanced diet. Because compound workouts are utilizing extra muscles so you
want to benefit from the strength positive aspects that come from that.
Features will differ from one individual to another depending on physique measurement and level of experience
in the health club.
Hypertrophy and power training are each kinds of resistance coaching.
Focusing on one area doesn’t imply you won’t see gains
within the other, but varying your workouts can optimize your results.
If your operational definition of "good" hinges on including bulk as shortly as potential, so you'll find a way to perform in a weight-dominant sport, a mass
gainer is strictly what you need. If, in distinction,
your goals are to maintain up an excellent physique
and add muscular energy, other sources of protein are
a greater bet.
Thus, indicating that a powerful bench does equate to an enormous chest in many
instances. Incline dumbbell presses put more emphasis on the clavicular head of the pecs and work several different
secondary muscular tissues as well. That Is due to the added shoulder flexion of
this movement. The primary reason for beginning with this exercise is to prioritize the upper chest.
This is usually the lagging portion of the chest in most people.
Nevertheless, I’ll additional focus on train order
in a while in this article. There are two forms of muscular hypertrophy, generally recognized as myofibrillar hypertrophy and sarcoplasmic hypertrophy.
That’s why we’ve laid out this simple and effective meal plan that can assist you placed on mass while staying lean. This is essential as a end
result of, with out correct restoration, you won’t make gains.
This shall be thought of a ‘push workout’ as you’re
working muscular tissues that push out. Some will choose
to coach triceps with chest, and that’s okay too (chest and triceps is
also a push workout). This will let you put your whole energy into constructing
large quads. There is no clear information to determine the type of protein, blend of amino acids,
and different vitamins that may make the best
supplement.
This article breaks down everything you want to know
when it comes to building muscle, including tips on how to work out,
what to eat, and restoration protocols. Diet and exercise work together and complement each other to improve muscle mass and
function. Pair this with our a la carte plan to fully customise your macros and meals selections, and make the dieting side
of muscle gain feel like a breeze. Ladies have a tendency to gain muscle in a quick time initially but can then gradual to a decrease price of features than males.
They are also much less prone to obtain a high muscle mass total.
The best way to gain mass for most people is through elevated energy and weight gain overall.
Barely higher in calorie than other meat sources, beef
is an efficient weight achieve meals as a end result of it is protein content material as nicely.
Whereas intake ought to be moderated, it can be a great addition here and there in your weight gain meal plan. Salmon incorporates
unsaturated fats that'll benefit your health as nicely as your calorie surplus.
One Other powerful advantage of intermittent fasting is
that it increases your circulating anabolic hormones,
corresponding to testosterone and progress hormone
(24). Both hormones help to build muscle and speed up recovery (25).
It’s recommended to take 5-10g of amino acids about half-hour earlier than a workout to prevent muscle fatigue,
stimulate protein synthesis, and assist velocity up muscle restoration.
Your muscular tissues grow when they’re resting and recovering, not when they’re being worked.
That’s why you should take loads of days off to ensure your muscle tissue are fully rested and recovered between lifts.
"If you've issues placing on muscle, your physique actually wants more time to recover than somebody who puts on muscle very easily," he says.
Four days within the gym with three days of
relaxation might simply do the trick. Research suggests that those
coaching to gain muscle should eat 1.4–2 grams of protein per
kg of body weight per day. To construct new muscle tissue, your main instruments
for rising your body’s price of protein synthesis are performing resistance training and getting sufficient quantities of
protein and overall vitamins. This process of accelerating your muscle mass is
named muscle hypertrophy, and it’s a main aim of resistance training.
Added muscle mass will increase the definition of your
muscular tissues, enhance your lean body mass, and add bulk and dimension to your frame in all the best places.
According to science, it appears men and women have a reasonably related response to resistance coaching.
Añadir un comentario
Fecha última actualización: 05/07/2021